

Drie kenmerken en functies in het organisme
- 838
- 195
- Irving McClure I
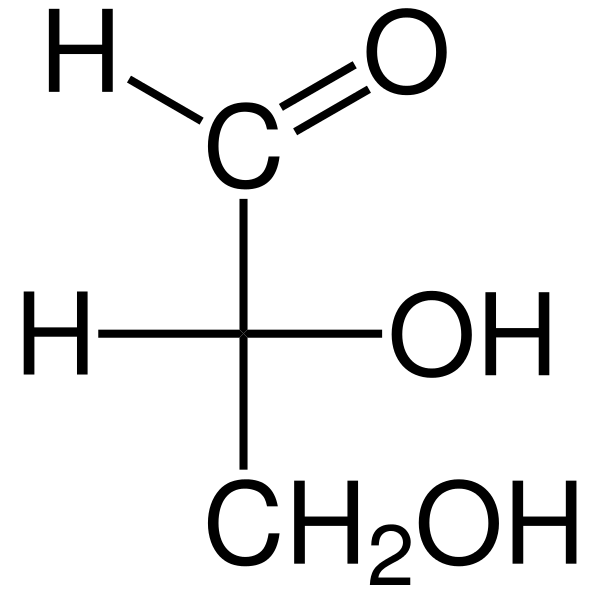
De Triosas Het zijn drie -koolstofmonosachariden waarvan de empirische chemische formule c is3H6OF6. Er zijn twee Triosas: GlyceralDehyd. De triosa's zijn belangrijk in het metabolisme omdat ze drie metabole routes verbinden: glycolyse, gluconeogenese en het pentosefosfaatpad.
Tijdens fotosynthese is de Calvin-cyclus een bron van triosas die dienen voor fructose-6-fosfaatbiosynthese. Deze suiker, op gefosforyleerde manier, wordt omgezet door enzymatisch gekatalyseerde stappen in reserve of structurele polysachariden.
Bron: Wesalius [Public Domain] De Triosas nemen deel aan de biosynthese van lipiden die deel uitmaken van celmembranen en adipocyten.
[TOC]
Kenmerken
De glyceraldehyde aldosa heeft een quiral koolstofatoom en heeft daarom twee enantiomeros, de l-glyceraldehyde en de d-glyceraldehyde. Zowel enantiomeren D en L hebben verschillende chemische en fysische kenmerken.
De D-glyceraldehyde verbrak het vlak van het gepolariseerde licht rechts (+) en heeft een rotatie [α] D, bij 25 ° C, van +8.7 °, terwijl het L-glyceraldehyde het vlak van het gepolariseerde licht aan de linkerkant (-) verbrak en een rotatie heeft [α] D, bij 25 ° C, van -8.7 °.
De chirale koolstof van glyceraldehyde is koolstof 2 (C-2), wat een secundaire alcohol is. De projectie van Fischer vertegenwoordigt de hydroxylgroep (-OH) van de d-glyceraldehyde rechts en de OH-groep van de linker L-Glyce.
Dihydroxyaceton mist chirale koolstofatomen en heeft geen enantiomere vormen. De toevoeging van een hydroxymethyleengroep (-choh) aan glyceraldehyde of dihydroxyaceton maakt het mogelijk om een nieuw chiraal centrum te creëren. Bijgevolg is suiker een tetrosa omdat het vier koolstofatomen heeft.
De toevoeging van een groep -chah aan Terosa creëert een nieuw chirale centrum. Gevormde suiker is een pentose. Groepen -Chah kunnen blijven toegevoegd aan een maximum van tien koolstofatomen.
Kan u van dienst zijn: fylogenieFuncties in het organisme
De triosa's als tussenpersonen in glycolyse, gluconeogenese en het pentosa -fosfaatpad
Glycolyse bestaat uit de breuk van het glucosemolecuul in twee pyruvaatmoleculen om energie te produceren. Deze route impliceert twee fasen: 1) voorbereidende fase of energieverbruik; 2) fase van energieopwekking. De eerste is degene die de Triosas produceert.
In de eerste fase wordt het glucosevrije energie -inhoud verhoogd, door de vorming van fosfoésters. In deze fase is het adenosine -triffosfaat (ATP) de fosfaatdonor. Deze fase culmineert met de omzetting van fructose fosfoéster 1,6-bifosfaat (F1.6BP) in twee triosas fosfaat, glyceraldehyde 3-fosfaat (GA3P) en dihydroxyaceton-fosfaat (DHAP).
Gluconeogenese is glucosebiosynthese van pyruvaat en andere tussenpersonen. Gebruik alle glycolyse -enzymen die reacties katalyseren waarvan de biochemische standaard energievariatie in evenwicht is (ΔGº '~ 0). Daarom hebben glycolyse en gluconeogenese gemeenschappelijke tussenpersonen, waaronder GA3P en DHAP.
Het pentosefosfaatpad bestaat uit twee stadia: een oxidatieve fase van glucose-6-fosfaat en een andere vorming van NADPH en ribosa-5-fosfaat. In de tweede fase wordt de 5-fosfaatribose omgezet in glycolyse-intermediairs, F1.6BP en GA3P.
De Triosas en de Calvin -cyclus
Fotosynthese is verdeeld in twee fasen. In de eerste treden licht -afhankelijke reacties op die NADPH en ATP produceren. Deze stoffen worden in de tweede gebruikt, waarbij er fixatie is van koolstofdioxide en hexosevorming uit triosas door een pad dat bekend staat als Calvin -cyclus.
In de Calvin-cyclus katalyseert de 1,5-biphary-oxygenase (Rubisco) ribulose-enzym de covalente unie van de CO2 Naar het pentose-ribulosa 1,5-bifosfaat en breekt de onstabiele intermediair van zes koolstofatomen in twee moleculen van drie koolstofatomen: de 3-fosfoglyceraat.
Kan u van dienst zijn: oligosachariden: kenmerken, compositie, functies, typenDoor enzymatische reacties die fosforylering en reductie van 3-fosfoglyceraat omvatten, optreedt met behulp van ATP en NADP, GA3P treedt op. Deze metaboliet wordt omgezet in 1,6-bifosfaatfructose (F1.6BP) door een metabole route vergelijkbaar met gluconeogenese.
Door werking van een fosfatase wordt F1.6BP omgezet in fructose-6-fosfaat. Vervolgens produceert een isomease-fosfhexose glucose 6-fosfaat (Glc6p). Ten slotte zet een epicherase GLC6P om in glucose 1-fosfaat, dat dient voor zetmeelbiosynthese.
De triosa's en lipiden van biologische en adipocytenmembranen
GA3P en DHAP kunnen fosfaatglycerol vormen, een noodzakelijke metaboliet voor de biosynthese van triacylglycerolen en glycerolipiden. Dit komt omdat beide triosa's fosfaat kunnen worden verbroken door een reactie die wordt gekatalyseerd door het Triosa -isomerase -fosfaat, dat beide Triosas in evenwicht houdt.
Het glycerol-fosfaat-enzymdehydrogenase katalyseert een oxidedringreactie, waarbij de NADH een paar elektronen aan de Dhap doneert om 3-fosfaatglycerol en NAD te vormen+. L-glycerol 3-fosfaat maakt deel uit van het skelet van fosfolipiden die een structureel deel uitmaken van biologische membranen.
De glycerol is proquiraal, het mist asymmetrische koolstofatomen, maar wanneer een van de twee primaire alcoholen een fosfoester vormt, kan het correct worden genoemd.
Glyceofosfolipiden worden ook fosfoglyceriden genoemd, worden genoemd als afgeleid van fosfatidinezuur. Fosfoglyceriden kunnen fosfoacylglycerolen vormen door esterbindingen te vormen met twee vetzuren. In dit geval is het resulterende product 1,2-fosfodiacylglycerol, een belangrijk onderdeel van membranen.
Een glyceofosfase katalyseert de hydrolyse van de 3-fosfaatglycerolfosfaatgroep, waardoor glycerol meer fosfaat produceert. Glycerol kan dienen als een startmetaboliet voor biosynthese van triacylglyceriden, die gebruikelijk zijn in adipocyten.
Kan u van dienst zijn: coagulase -test: fundering, procedure en gebruikDe Triosas en Membranen van de Archeobacteria
Vergelijkbaar met eubacteriën en eukaryoten, wordt 3-fosfaatglycerol gevormd uit Triosas-fosfaat (GA3P en DHAP). Er zijn echter verschillen: de eerste is dat 3-fosfaatglycerol in archeobacteria membranen L-configuratie is, terwijl het in de eubacteriën en eukaryotische membranen configuratie D is.
Een tweede verschil is dat archeobacteria membranen ester vormen met twee lange koolwaterstofketens van isoprenoïde groepen, terwijl in eubacteriën en eukaryoten de glycerol esterbindingen (1,2-dicilglycerol) vormt met twee koolwaterstofketens van vetzuren van vetzuren van vetzuren van vetzuren van vetzuren en eubacterie.
Een derde verschil is dat, in archeobacteria membranen, vervangingen in het fosfaat en 3-fosfaatglycerol verschillen van die van eubacteriën en eukaryoten. De fosfaatgroep is bijvoorbeeld gekoppeld aan disaccharide α-Glucopiranso- (1®2)-β-Galatofuranosa.
Referenties
- Cui, s. W. 2005. Voedselkoolhydraten: chemie, fysische eigenschappen en toepassingen. CRC Press, Boca Raton.
- Pik, p., Mäkinen, K, Honkala, en., Saag, m., Kennepohl, E., EAPEN, A. 2016. Erythritol is effectiever dan xylitol en sorbitol bij het beheren van eindpunten van orale gezondheid. International Journal of Dentistry.
- Nelson, D. L., Cox, m. M. 2017. Lehninger -principes van biochemie. W. H. Freeman, New York.
- Sinnott, m. L. 2007. Koolhydraatchemie en biochemie structuur en mechanisme. Royal Society of Chemistry, Cambridge.
- Stick, r. V., Williams, s. J. 2009. Koolhydraten: de essentiële moleculen van het leven. Elsevier, Amsterdam.
- VOET, D., VOET, J. G., Pratt, c. W. 2008. Fundamentals of Biochemistry - Life op moleculair niveau. Wiley, Hoboken.
- « Aldose -kenmerken, typen, aantal koolstofatomen
- Fluoxetine (Prozac) werkingsmechanisme en bijwerkingen »