

Synthese van vetzuren waar het gebeurt, enzymen, stadia en reacties
- 3100
- 950
- Dewey Powlowski
De vetzuursynthese Het is het proces waarbij de fundamentele componenten van de belangrijkste lipiden van de cellen (vetzuren), die deelnemen aan veel zeer relevante cellulaire functies worden geproduceerd.
Vetzuren zijn alifatische moleculen, dat wil zeggen dat ze in wezen zijn samengesteld uit koolstof- en waterstofatomen die door elkaar zijn verenigd, min of meer lineair. Ze hebben een methylgroep aan een van hun terminale uiteinden en een zuurcarbonylische groep in de andere, waarvoor ze "vetzuren" worden genoemd.
Samenvatting van de synthese van vetzuren (Bron: Mephist.Org/licenties/by-sa/3.0) Via Wikimedia Commons) Lipiden zijn moleculen die worden gebruikt door verschillende celbiosynthetische systemen voor de vorming van andere meer complexe moleculen zoals:
- membraanfosfolipiden
- triglyceriden voor energieopslag en
- De ankers van enkele speciale moleculen gevonden op het oppervlak van vele soorten cellen (eukaryoten en prokaryoten)
Deze verbindingen kunnen bestaan als lineaire moleculen (met alle koolstofatomen verzadigd met waterstofmoleculen), maar die van lineaire keten kunnen ook worden waargenomen en met sommige verzadigingen, dat wil zeggen met dubbele bindingen tussen de koolstofatomen.
Verzadigde vetzuren kunnen ook vertakte ketens vinden, waarvan de structuur een beetje complexer is.
De moleculaire kenmerken van vetzuren zijn cruciaal voor hun functie, omdat velen afhankelijk zijn van de fysicochemische eigenschappen van de moleculen die hierdoor worden gevormd, met name hun smeltpunt, hun mate van verpakking en hun capaciteit voor bicapas -formatie.
Vetzuursynthese is dus een extreem gereguleerde materie, omdat het een reeks kritische sequentiële gebeurtenissen is voor de cel vanuit vele gezichtspunten.
[TOC]
Waar gebeurt de synthese van vetzuren??
Bij de meeste levende organismen vindt de synthese van vetzuren plaats in het cytosolische compartiment, terwijl hun afbraak voornamelijk optreedt tussen cytosol en mitochondria.
Het proces hangt af van de energie in de ATP-bindingen, het NADPH-verminderingsvermogen (meestal afgeleid van de penty fosfaatroute), de biotine-cofactor, bicarbonaationen (HCO3-) en mangaanionenionen ionen.
Bij zoogdieren zijn de belangrijkste organen van vetzuursynthese de lever, nieren, hersenen, longen, borstklieren en vetweefsel.
Het directe substraat van synthese van novo Van vetzuren is acetyl-CoA en het eindproduct is een palmitaatmolecuul.
Kan u van dienst zijn: BHI Agar: wat is, fundering, voorbereiding, gebruikAcetyl-CoA komt rechtstreeks voort uit de verwerking van glucolithische intermediairs, en daarom bevordert een dieet met koolhydraten de synthese van lipiden (lipogenese) ergo, ook van vetzuren.
Enzymen die deelnemen
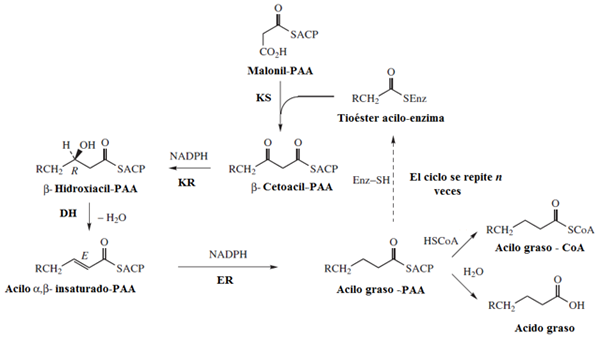
Acetyl-CoA is het twee-koolstofsyntheseblok dat wordt gebruikt voor de vorming van vetzuren, omdat verschillende van deze moleculen opeenvolgend worden verbonden met een malonyl-CoA-molecuul, gevormd door de carboxylering van een acetyl-CoA.
Het eerste enzym van de route, en een van de belangrijkste vanuit het oogpunt van de regulering, is die persoon die verantwoordelijk is voor de carboxylering van acetyl-CoA, bekend als acetyl-CoA-carboxylase (ACC), wat een complex enzymatisch is gevormd door 4 eiwitten en gebruikt biotine als een cofactor.
En ondanks het feit dat er structurele verschillen zijn tussen verschillende soorten, is het enzymvetzuursynthase degene die de leiding heeft over de belangrijkste biosynthetische reacties.
Dit enzym is in werkelijkheid een enzymatisch complex bestaande uit monomeren met de 7 verschillende enzymatische activiteiten, die nodig zijn voor de rek van vetzuur in "geboorte".
De 7 activiteiten van dit enzym kunnen als volgt worden vermeld:
- ACP: Acilo Group Transporteer eiwit
- Acetyl-CoA-ACP transacetilasa (BIJ)
- β-cetoacil-ACP-synthase (KS)
- Malonyl-CoA-ACP-transferase (MT)
- β-cethoacil-ACP-reductase (KR)
- β-hydroxyacil-ACP dehydratase (HD)
- Enoil-ACP Redtase (Eh)
In sommige organismen zoals bacteriën, bijvoorbeeld, wordt het synthase -vetzuurcomplex gevormd door onafhankelijke eiwitten die met elkaar worden geassocieerd, maar worden gecodeerd door verschillende genen (synthase type II van vetzuursystemen).
 Vetzuursynthesa van gist (bron: xiong, en., Lomakin, ik.B., Steitz, T.NAAR. / Public Domain, via Wikimedia Commons)
Vetzuursynthesa van gist (bron: xiong, en., Lomakin, ik.B., Steitz, T.NAAR. / Public Domain, via Wikimedia Commons) In veel eukaryoten en sommige bacteriën bevat het multienzym echter verschillende katalytische activiteiten die worden gescheiden in verschillende functionele domeinen, in een of meer polypeptiden, maar die kunnen worden gecodeerd door hetzelfde gen (synthase type I) van het vetzuursysteem).
Fasen en reacties
De meeste onderzoeken die zijn uitgevoerd met betrekking tot de synthese van vetzuren omvatten de bevindingen die in het bacteriële model zijn gedaan, maar de synthese -mechanismen van eukaryotische organismen zijn ook met enige diepte bestudeerd.
Het is belangrijk om te vermelden dat het vetzuursysteem type II wordt gekenmerkt dat alle vetacyl intermediairs covalent een klein groot zuur eiwit verbinden dat bekend staat als het acyltransporteiwit (ACP), dat ze van de ene enzym naar het volgende transporteert.
Kan u van dienst zijn: ABO System: onverenigbaarheid, erfenis en bewijsIn eukaryoten maakt ACP -activiteit integendeel deel uit van hetzelfde molecuul, waardoor het begrijpt dat enzym zelf een speciale plaats heeft voor de vereniging van intermediairs en hun transport door de verschillende katalytische domeinen.
De vereniging tussen het eiwit of het ACP -gedeelte en de vette acil.
- Aanvankelijk is het acetyl-CoA-carboxylase (ACC) enzym verantwoordelijk voor het katalyseren van de eerste stap van "commitment" in de synthese van vetzuren die, zoals vermeld, de carboxylering van een acetyl-CoA-molecuul impliceert om de tussenpersoon van 3 koolstofatomen bekend te maken Als malonyl-coa.
Het synthase -vetzuurcomplex ontvangt acetyl- en malonil -groepen, die de "tiol" -plaatsen hiervan correct moeten vullen.
Dit vindt aanvankelijk plaats voor de overdracht van acetyl-CoA naar de Cisteína SH-groep in de β-zo-beacil-ACP-synthase, reactie gekatalyseerd door acetyl-CoA-ACP transacetilas.
De malonylgroep wordt overgebracht van malonyl-CoA naar de SH-groep van het ACP-eiwit, een gebeurtenis gemedieerd door het malonyl-CoA-ACP-transferase-enzym, dat malonyl-ACP vormt.
- De initiatie van de verlenging van vetzuur bij de geboorte bestaat uit de condensatie van de malonil-ACP met een acetyl-CoA-molecuul, een reactie gericht door een enzym met β-cethoacyl-ACP-activiteitssynthase. In deze reactie wordt acetoacethyl-ACP gevormd en wordt een CO2-molecuul vrijgegeven.
- Verlengingsreacties treden op in cycli waarbij 2 koolstofatomen tegelijkertijd worden toegevoegd, dat elke cyclus bestaat uit een condensatie, een reductie, een uitdroging en een tweede reductie -gebeurtenis:
- Condensatie: Acetyl- en malonylgroepen zijn gecondenseerd om acetoacetyl-ACP te vormen
- Reductie van de carbonylgroep: de koolstofgroep van koolstof 3 van de acetoacetyl-ACP is verminderd en vormt D-β-hydroxybuteril-ACP, reactie gekatalyseerd door de β-cethoacyl-ACP-Reductase, die Nadph gebruikt als elektronendonor.
- Uitdroging: hydrogenen tussen koolstofatomen 2 en 3 van het vorige molecuul worden verwijderd, waardoor een dubbele binding wordt gevormd die eindigt met de productie van de trans-∆2-butenoil-ACP. De reactie wordt gekatalyseerd door β-hydroxyacil-ACP dehydratase.
- Dubbele linkreductie: de dubbele binding van trans-∆2-buTenoil-ACP wordt gereduceerd tot Butiril-ACP door werking van de woede-ACP-reductase, die NADPH ook gebruikt als een reductiemiddel.
Het kan u van dienst zijn: Chiapas Flora en Fauna: representatieve soortenOm door te gaan met de verlenging, moet een nieuw malonylmolecuul zich aansluiten.
 Palmitato Structure (Bron: Edgar181 / Public Domain, via Wikimedia Commons)
Palmitato Structure (Bron: Edgar181 / Public Domain, via Wikimedia Commons) In elke verlengingsstap wordt een nieuw malonyl-CoA-molecuul gebruikt om de keten te laten groeien in 2 koolstofatomen en deze reacties worden herhaald totdat de juiste lengte is bereikt (16 koolstofatomen), waarna een enzym liberase thioesterase vol vetzuur door hydratatie.
Palmitaat kan vervolgens worden verwerkt door verschillende soorten enzymen die hun chemische kenmerken wijzigen, dat wil zeggen dat ze onverzadiging kunnen introduceren, hun lengte kunnen verlengen, enz.
Regulatie
Zoals veel biosynthetische of afbraakroutes, wordt vetzuursynthese gereguleerd door verschillende factoren:
- Het hangt af van de aanwezigheid van bicarbonaationen (HCO3-), van vitamine B (biotine) en acetyl-CoA (tijdens de initiële doorgang van de route, die de carboxylering van een acetyl-CoA-molecuul impliceert door middel van een carboxyard tussenpersoon van biotine om malonyl-coa te vormen).
- Het is een route die optreedt als reactie op de kenmerken van cellulaire energie, omdat wanneer er voldoende hoeveelheid "metabole brandstof" is, overmaat wordt omgezet in vetzuren die worden opgeslagen voor latere oxidatie in momenten van energietekort.
In termen van de regulatie van het acetyl-CoA-carboxylase-enzym, dat de beperkende stap van de gehele route vertegenwoordigt, wordt dit geremd door de palmitail-CoA, het belangrijkste product van de synthese.
Zijn toostale activator daarentegen is citraat, dat het metabolisme leidt van oxidatie naar zijn synthese voor opslag.
Wanneer acetyl-CoA en ATP mitochondriale concentraties toenemen, wordt citraat getransporteerd naar cytosol, waar het zo voorloper is voor acetyl-CoA-cytosolische synthese en een alkalistisch activeringssignaal voor acetyl-CoA-carboxylase.
Dit enzym kan ook worden gereguleerd door fosforylering, gebeurtenis afgevuurd door de hormonale werking van glucagon en epinefrine.
Referenties
- McGenity, T., Van der Meer, j. R., & De Lorenzo, V. (2010). Handboek van koolwaterstof- en lipidenmicrobiologie (P. 4716). K. N. Timmis (ed.)). Berlijn: Springer.
- Murray, r. K., Granner, D. K., Mayes, p. NAAR., & Rodwell, V. W. (2014). Harper's geïllustreerde biochemie. McGraw-Hill.
- Nelson, D. L., & Cox, m. M. (2009). Lehninger -principes van biochemie (pp. 71-85). New York: WH Freeman.
- Numa, s. (1984). Vetzuurmetabolisme en zijn regulering. Elsevier.
- Rawn, J. D. (1989). Biochemie-internationale editie. North Carolina: Neil Patterson Publishers, 5.
- « Systematische fout hoe het te berekenen, in chemie, in fysica, voorbeelden
- Onverzadigde vetzurenkenmerken, structuur, functies, voorbeelden »