

Lipogenese -kenmerken, functies en reacties
- 4343
- 424
- Cecil Graham
De lipogenese Het is de belangrijkste metabole route waardoor vetzuren lange ketens worden gesynthetiseerd door overtollige koolhydraten in het dieet. Deze vetzuren kunnen worden opgenomen in triglyceriden door middel van sterificatie tot glycerolmoleculen.
Onder normale omstandigheden vindt lipogenese plaats in de lever en vetweefsel en wordt beschouwd als een van de belangrijkste belastingbetalers van het onderhoud van triglyceridische homeostase in bloedserum.
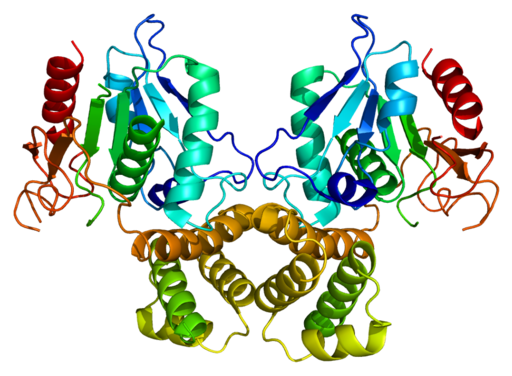
 Human Syntasa Fat Structure (FASN) (Bron: EMW
Human Syntasa Fat Structure (FASN) (Bron: EMW[CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via
Wikimedia Commons)
Triglyceriden zijn het belangrijkste energiereservoir van het lichaam en de energie hierin wordt geëxtraheerd dankzij een proces dat bekend staat als lipolyse dat, in tegenstelling tot lipogenese, bestaat uit de scheiding en afgifte van glycerolmoleculen en vetzuren in de richting.
De vrijgegeven glycerol dient als een substraat voor de gluconeogene route en vetzuren kunnen worden getransporteerd naar andere gratis complementaire compartimenten met serumalbumine.
Deze vetzuren worden vastgelegd door bijna alle weefsels met uitzondering van de hersenen en erytrocyten, dan worden ze opnieuw gesterifieerd tot triacylglycerol om te worden geoxideerd als brandstoffen of opgeslagen als een energiereserve.
Vet -rijke diëten zijn de belangrijkste oorzaken van obesitas, omdat het calorie -overtollige moet worden opgeslagen en vetweefsel moet uitzetten om zowel overtollig -ingesteste lipiden als die en endogeen te accommoderen.
[TOC]
Kenmerken en functies
In het menselijk lichaam ontstaan bijvoorbeeld vetzuren van biosynthetische processen van acetyl-CoA of als een product van de hydrolytische verwerking van membraanvet en fosfolipiden.
Veel zoogdieren zijn niet in staat om sommige vetzuren te synthetiseren, wat deze essentiële componenten van hun dieet maakt.
De belangrijkste functie van lipogenese heeft te maken met de opslag van energie in de vorm van vet (lipiden) die optreedt bij het consumeren van een grotere hoeveelheid koolhydraten dan die die het lichaam nodig heeft, zelfs de hepatische opslagcapaciteiten van glycogeen overschrijden.
Kan u bedienen: Gastrine: kenmerken, structuur, productie, functiesLipiden gesynthetiseerd door deze route worden opgeslagen in het witte vetweefsel, de belangrijkste plaats van opslag van lipiden in het lichaam.
In alle lichaamscellen treedt lipogenese echter op, echter, vetweefsels en de lever zijn de belangrijkste plaatsen van synthese. Deze route wordt gegeven in het celcytoplasma, terwijl de oxidatie van vetzuren optreedt in mitochondriale compartimenten.
Lipogenese en daaropvolgende synthese van triglyceriden wordt gevolgd door de synthese en secretie van lipoproteische deeltjes met zeer lage dichtheid bekend als VLDL -deeltjes (Engels Variëren lipoproteïne met lage dichtheid), die in staat zijn om de bloedbaan binnen te gaan.
Zowel VLDL -deeltjes als triglyceride.
Reacties
De stroom van koolstofatomen van de glucose die aanwezig is in koolhydraten tot vetzuren wordt gemoduleerd door lipogenese en omvat een reeks perfect gecoördineerde enzymatische reacties.
1-De glycolytische route in de cytosol van de cellen is verantwoordelijk voor het verwerken van de glucose die vanuit de bloedbaan binnenkomt om pyruvaat te produceren, dat wordt omgezet in acetyl-CoA, in staat om de Krebs-cyclus in de mitochondria in te voeren, waar citraten optreedt, waar citraten optreedt, waar citraten optreedt, waar citraten optreedt.
2-De eerste stap van de lipogene route bestaat uit de conversie van citraat die de mitochondria in acetyl-CoA verlaat door de werking van een enzym dat bekend staat als ATP-citrate liasa (ACYY).
3-Het resulterende acetyl-CoA is gearboxyleerd om malonyl-CoA te vormen, een acetyl-CoA-carboxylase (aqueca).
4-De derde reactie is de reactie die de beperkende stap van de gehele route oplegt, dat wil zeggen de langzaamste reactie, en bestaat uit de conversie van de malonyl-CoA om te palmiteren door een enzymvetzuursynthase (FAS) (FAS) (FAS).
Het kan u van dienst zijn: Half Cary Blair: fundering, voorbereiding en gebruik5-een andere stroomafwaartse reacties helpen het palmitaat om te zetten in andere meer complexe vetzuren, maar het palmitaat is het belangrijkste product van lipogenese van novo.
Vetzuursynthese
De synthese van vetzuren bij zoogdieren begint met de complexe vetzuursynthase (FAS), een multifunctioneel en multi -verstopend complex in het cytosol dat het palmitaat synthetiseert (een 16 koolstof verzadigd vetzuur). Voor deze reactie gebruikt hij, zoals al vermeld, malonyl-Coa als koolstofdonor en NADPH als een cofactor.
De subeenheden van de homodimer van de FAS katalyseren de synthese en verlenging van vetzuren twee koolstofatomen tegelijkertijd. Deze subeenheden hebben zes verschillende enzymatische activiteiten: acetyltransferase, b-cetoacil syntasa, malonyl transferas.
Verschillende leden van een familie van verlengingseiwit van zeer lange ketenvetzuren (ELOVL) zijn verantwoordelijk voor het verlengen van de vetzuren die door de FAS worden geproduceerd. Stroomafwaarts zijn andere enzymen die verantwoordelijk zijn voor de introductie van dubbele bindingen (desaturatie) in de ketens van vetzuren.
Regulatie
Talrijke pathofysiologische omstandigheden hebben te maken met de defecte regulatie van de lipogene route, aangezien onregelmatigheden in dezelfde lipide homeostase met onderbrekingen.
Een dieet dat rijk is aan koolhydraten activeert lipogenese, maar er is aangetoond dat het niet alleen de hoeveelheid koolhydraten is, maar ook het type koolhydraten.
Experimentele gegevens tonen bijvoorbeeld aan dat eenvoudige suikers zoals fructose veel krachtigere effecten hebben op de activering van lipogenese dan andere complexere koolhydraten.
Glucolithisch glucosemetabolisme vertegenwoordigt een grote bron van koolstof voor vetzuursynthese.
Glucose induceert ook de expressie van de enzymen die betrokken zijn bij de lipogene route door vakbondseiwitten naar koolhydraatresponselementen.
Kan u dienen: San Luis Potosí Flora en Fauna: meer representatieve soortenBloedglucosespiegels stimuleren ook de expressie van deze enzymen door stimulatie van insulinebevrijding en remming van glucagonbevrijding in de pancreas. Dit effect wordt geregeld via het eiwit van binding aan het regulerende element van sterol 1 (SREBP-1) in levercellen en adipocyten.
Andere regulatieroutes hebben veel te maken met het endocriene systeem en verschillende hormonen indirect gerelateerd aan de expressie van veel van de lipogene enzymen.
Referenties
- Ameer, f., Scandiuzzi, L., Hasnain, s., Kalbacher, h., & Zaidi, n. (2014). Van novo lipogenese in gezondheid en ziekte. Metabolisme, 0-7.
- Lodhi, ik. J., Wei, x., & Semenkovich, c. F. (2011). Lipoexpedy: van novo lipogenese als een metabolische signaalzender. Trends in endocrinologie en metabolisme, 22(1), 1-8.
- Mathews, c., Van Holde, K., & Ahern, k. (2000). Biochemie (3e ed.)). San Francisco, Californië: Pearson.
- Nelson, D. L., & Cox, m. M. (2009). Lehninger -principes van biochemie. Omega -edities (5e ed.)).
- Samuel, v. T. (2011). Fructose geïnduceerde lipogenese: van suiker tot vet tot insulineresistentie. Trends in endocrinologie en metabolisme, 22(2), 60-65.
- Scherer, T., Hare, J. OF., Diggs-Andrews, K., Schweiger, m., Cheng, B., Lindtner, C.,... Buettner, c. (2011). Hersensuline regelt vetweefsellipolyse en lipogenese. Celmetabolisme, 13(2), 183-194.
- Schutz, en. (2004). Dieetvet, lipogenese en energiebalans. Fysiologie en gedrag, 83, 557-564.
- Stabiel, m. S., & Ntambi, j. M. (2010). Genetische controle van novo-lipogenese: rol bij door dieet geïnduceerde obesitas. Kritische beoordelingen in biochemie en moleculaire biologie, Vier vijf(3), 199-214.
- Zaidi, n., Lupien, l., Kuemmerle, n. B., Kinlaw, w. B., Swinnen, J. V, & Smans, K. (2013). Lipogenese en lipolyse: de paden die door de kankercellen worden uitgebuit om vetzuren vetzuren te verwerven. Vooruitgang in lipide -onderzoek, 52(4), 585-589.
- « Actieve leerkenmerken, stijlen en strategieën
- Moderne filosofische kenmerken, fasen en vertegenwoordigers »