

Glycogeenstructuur, synthese, afbraak, functies
- 3724
- 350
- Miss Herman Russel
Hij Glycogeen Het is de opslagkoolhydraten van de meeste zoogdieren. Koolhydraten worden vaak suikers genoemd en deze worden geclassificeerd volgens het aantal afval dat wordt veroorzaakt door hydrolyse (monosachariden, disacchariden, oligosachariden en polysachariden)))).
Monosachariden zijn de eenvoudigste koolhydraten die zijn geclassificeerd volgens het aantal koolstofatomen in hun structuur. Er zijn dan de Triosas (3C), Tetrosas (4C), Pentosas (5C), hexosous (6C), heptosase (7C) en octosa's (8c).
Chemische glycogeenstructuur met glycosidebindingen (bron: glykogeen.SVG: Neurotoger Derivative Work: Marek M [Public Domain] via Wikimedia Commons) Volgens de aanwezigheid van de Aldehyde -groep of de Cetona -groep zijn deze monosachariden ook geclassificeerd als respectievelijk aldies of ketosas.
Disacchariden geven aanleiding tot hydrolyse, twee eenvoudige monosachariden, terwijl oligosachariden 2 tot 10 eenheden monosachariden en polysachariden produceren, produceren meer dan 10 monosachariden.
Het glycogeen is vanuit biochemisch oogpunt een polysacharide bestaande uit vertakte ketens van een zes -koolstofaldose, dat wil zeggen een hexose bekend als glucose. Grafisch kan het worden weergegeven voor glycogeen als een glucoseboom. Dit wordt ook wel dierenzetmeel genoemd.
Glucose in planten wordt opgeslagen als zetmeel en bij dieren als glycogeen, dat voornamelijk wordt bewaard in de lever- en spierweefsel.
In de lever kan glycogeen 10% van zijn massa en 1% spiermassa vestigen. Net als bij een man van 70 kg weegt de lever ongeveer 1800 g en de spieren ongeveer 35 kg, de totale hoeveelheid spierglycogeen is veel groter dan lever.
[TOC]
Structuur
Het molecuulgewicht van glycogeen kan 108 g/mol bereiken, gelijkwaardig aan 6 x 105 glucosemoleculen. Glycogeen bestaat uit meerdere vertakte a-D-glycoseketens. Glucose (C6H12O6) is een aldohexosa die op een lineaire of cyclische manier kan worden weergegeven.
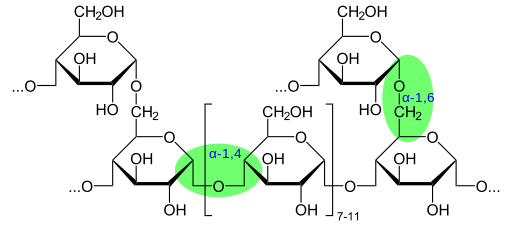
De glycogeen heeft een zeer vertakte en compacte structuur met ketens van 12 tot 14 glucoseafval in de vorm van α-D-glucose die zijn gekoppeld aan α- (1 → 4) glucosidebindingen. De ketting vertakkingen worden gevormd door α- (1 → 6) glucosidische links.
Glycogeen, zoals het zetmeel dat in het dieet wordt ingenomen, biedt de meeste koolhydraten die het lichaam nodig heeft. In de darm worden deze polysachariden afgebroken door hydrolyse en vervolgens geabsorbeerd naar de bloedsomloopstroom, voornamelijk als glucose.
Drie enzymen: ß-amylase, a-amylase en amylo-a- (1 → 6) -glucosidase zijn verantwoordelijk voor de darmafbraak van zowel glycogeen als zetmeel.
Het a-amylase hydrolyseert willekeurig de α-bindingen (1 → 4) van de zijketens van zowel glycogeen als zetmeel, en ontvangt daarom de naam van endoglysidase. De ß-AMYLA is een exoglicosidase die ß-maltose dímeros vrijgeeft die α- breekt (1 → 4) glycosidische schakels vanaf de uiteinden van de buitenste ketens zonder de gevolgen te bereiken.
Gezien het feit dat noch ß-amylase noch α-amylase takken afbreken, is het eindproduct van zijn werking een zeer vertakte structuur van ongeveer 35 tot 40 glucoseresten die de limiet dextrine worden genoemd.
De limietdextrine wordt uiteindelijk gehydrolyseerd op de vertakkingspunten die α- (1 → 6) bindt door de amyle-a- (1 → 6) -glucosidase, ook bekend als "lasterlijk" enzym. De ketens die worden vrijgegeven door deze defloat worden na afgebroken door ß-amylase en a-amylase.
Terwijl het ingenomen glycogeen binnenkomt als glucose, moet degene die in de weefsels wordt gevonden, door het organisme worden gesynthetiseerd door glucose.
Kan u van dienst zijn: Purines: kenmerken, structuur, functiesSynthese
Glycogeensynthese wordt glycogenese genoemd en vindt vooral plaats in de spier en lever. De glucose die het organisme binnengaat met het dieet gaat door naar de bloedsomloopstroom en van daaruit in de cellen, waar het onmiddellijk wordt gefosforyleerd door een enzym genaamd glycquinase.
Glucoquinase fosforyyl tot glucose bij koolstof 6. De ATP biedt fosfor en energie voor deze reactie. Als gevolg hiervan wordt glucose 6-fosfaat gevormd en wordt een ADP vrijgegeven. Dan wordt 6-fosfaatglucose glucose 1-fosfaat door de werking van een fosfoglucomutase die de fosfor van positie 6 naar positie 1 moddert 1.
De 1-fosfaatglucose wordt geactiveerd voor glycogeensynthese, wat de deelname van een set van drie andere enzymen impliceert: de UDP-glycose pyrofosforylase, synthetische glycogeen en amilo- (1,4 → 1.6) -glicosferase.
Glucose-1-fosfaat, samen met de trifosfaat-uridine (UTP, een nucleoside van uridine trifosfaat) en door werking van het UDP-glycose-pyrofosforylase, vormen het difosfaat-glucose uridinecomplex (UDP GLC) (UDP GLC) (UDP GLC) (UDP GLC) (UDP GLC). In het proces wordt een pyrofosfaation gehydrolyseerd.
Vervolgens vormt het gesynthetteerde glycogeen enzym een glucosidebinding tussen het C1 van het UDP GLC -complex en de C4 van een glycogeen glucose -terminaalresidu en de UDP UDP komt vrij uit het UDP -geactiveerde glucosecomplex. Om deze reactie te laten optreden, moet er een pre -bestaande glycogeenmolecule zijn die "primaire glycogeen" wordt genoemd.
Het primordiale glycogeen wordt gesynthetiseerd op een priming -eiwit, glycogenine, dat 37 kDa en Glysila heeft in een tyrosineresten met behulp van het UDP GLC -complex. Van daaruit zijn ze gekoppeld α-D-glucoseafval met 1 → 4 verbindingen en wordt een kleine ketting gevormd waarop de synthesase-glycogeen werkt.
Zodra de initiële ketting ten minste 11 glucoseresten verbindt, draagt het vertakking of amile -enzym (1,4 → 1,6) -glicosyltransferase een kettingstuk van 6 of 7 glucoseverval over aan de aangrenzende keten in positie 1 → 6, die een tak vestigt punt. Het aldus gebouwde glycogeenmolecuul wordt groeit door toevoegingen van glucose -eenheden met glycosidische links 1 → 4 en meer gevolgen.
Degradatie
Glycogeenafbraak wordt glucogenolyse genoemd en is niet equivalent aan het omgekeerde pad van de synthese. De snelheid van deze route wordt beperkt door de snelheid van de reactie die wordt gekatalyseerd door de fosforylase glycogeen.
Fosforyllase glycogeen is verantwoordelijk voor het splitsen (fosforolyse) van koppelingen 1 → 4 uit glycogeenketens, waardoor glucose 1-fosfaat vrijgeeft. De enzymatische werking begint aan de uiteinden van de buitenste ketens en worden achtereenvolgens verwijderd totdat 4 glucoseresten aan elke kant van de gevolgen blijven.
Vervolgens, een ander enzym, het α- (1 → 4) → α- (1 → 4) glucano-transferas, laat het vertakkingspunt blootgesteld door een trisaccharide-eenheid van de ene tak naar de andere over te brengen. Hierdoor kan de amilo- (1 → 6) -glucosidase (niet -rampische enzym) hydrolys mogelijk zijn. De gecombineerde werking van deze enzymen eindigt volledig op splijten naar glycogeen.
Aangezien de initiële reactie van fosfomutase omkeerbaar is, kan 6-fosfaatglucose worden gevormd uit glucoseresten 1-fosfaat gesplitst uit glycogeen. In de lever en nier, maar niet in de spier, is er een enzym, glucose-6-fosfatase, in staat om te verzamelen tot 6-fosfaatglucose en er in vrije glucose om te veranderen.
Kan u van dienst zijn: FOTOLYSEDefosforyleerde glucose kan zich tot bloed verspreiden, en dat is hoe hepatische glycogenolyse wordt weerspiegeld in een toename van de bloedglucosewaarden (glycemie).
Regulatie van synthese en afbraak
Synthese
Dit proces wordt uitgeoefend op twee fundamentele enzymen: synthesase glycogeen en fosforylase glycogeen, zodat wanneer een van hen wordt geactiveerd, de andere zich in zijn inactieve toestand bevindt. Deze regulatie voorkomt tegengestelde reacties van synthese en afbraak dat tegelijkertijd plaatsvindt.
De actieve vorm en inactieve vorm van beide enzymen is zeer verschillen.
Adrenaline is een hormoon dat wordt vrijgelaten uit het bijniermerg, en glucagon is een andere die voorkomt in het endocriene deel van de alvleesklier. Endocriene pancreas produceert insuline en glucagon. Langerhans eilandjes α zijn degenen die de glucagon synthetiseren.
Adrenaline en glucagon zijn twee hormonen die worden vrijgegeven wanneer energie nodig is als reactie op de afname van de bloedglucosespiegels. Deze hormonen stimuleren de activering van fosforylase glycogeen en remmen synthesase -glycogeen, waardoor glycogenolyse wordt gestimuleerd en glycogenese remmen.
Terwijl adrenaline zijn werking op spier en lever uitoefent, werkt Glucagon alleen op de lever. Deze hormonen zijn verbonden met specifieke membranale receptoren in de witte cel, die cyclasa -adenilaat activeert.
De activering van het cyclase -adenylaat begint een enzymatische waterval die enerzijds een AMPC -afhankelijk eiwitquinase activeert dat inactief is voor synthetische glycogeen en activeert glycogeenfosforylase door fosforylering (respectievelijk direct en indirect,).
De skeletspier heeft een ander mechanisme voor activering van fosforylase glycogeen door calcium, dat wordt afgegeven als een gevolg van depolarisatie van het spiermembraan aan het begin van de samentrekking.
Degradatie
Enzymatische watervallen die hierboven zijn beschreven, verhogen uiteindelijk de glucosewaarden en wanneer ze een bepaald niveau bereiken, wordt glycogenese geactiveerd en wordt glucogenolyse geremd, waardoor ook de verdere afgifte van adrenaline en glucagon wordt geremd.
Glycogenese wordt geactiveerd door activering van fosfatase fosforylase, een enzym dat glycogeensynthese reguleert door verschillende mechanismen, die de inactivering van het kinase -fosforylase en fosforylase α impliceren, dat een synthesase -glycogeenremmer is.
Insuline bevordert de binnenkomst van glucose in spiercellen, waardoor 6-fosfaatglucosespiegels worden verhoogd, wat defosforylering en activering van synthesase-glycogeen stimuleert. Aldus begint de synthese en wordt de afbraak van glycogeen geremd.
Functie
Spierglycogeen vormt een energiereserve voor de spier die, net als reservevetten, spieren toestaat om zijn functies te vervullen. Omdat het een glucosebron is, wordt spierglycogeen gebruikt tijdens het sporten. Deze reserveringen nemen toe met fysieke training.
In de lever vormt glycogeen ook een belangrijke reservebron voor zowel de functies van het orgaan als voor de bijdrage van glucose aan de rest van het lichaam.
Deze functie van leverglycogeen is te wijten aan het feit dat de lever 6-fosfatase-glucose bevat, een enzym dat in staat is de 6-fosfaatglucosefosfaatgroep te elimineren en deze in vrije glucose te maken. Vrije glucose kan, in tegenstelling tot gefosforyleerde glucose, worden verspreid door het hepatocytenmembraan (levercellen).
Het kan u van dienst zijn: sporulatie: in planten, in schimmels en in bacteriënDit is hoe de lever glucose naar circulatie kan bieden en stabiele glucosespiegels kan behouden, zelfs in langdurige vastencondities.
Deze functie is van groot belang, omdat de hersenen bijna uitsluitend worden gevoed door bloedglucose, kan ernstige hypoglykemie (zeer lage bloedglucoseconcentraties) verlies van kennis veroorzaken.
Gerelateerde ziekten
Glycogeen -gerelateerde ziekten ontvangen de generieke naam van "glycogeenopslagziekten".
Deze ziekten vormen een groep erfelijke pathologieën die worden gekenmerkt door de afzetting in de weefsels van abnormale hoeveelheden of soorten glycogeen.
De meeste glycogeenopslagziekten worden veroorzaakt door een genetisch aardtekort van een van de enzymen die betrokken zijn bij glycogeenmetabolisme.
Ze zijn ingedeeld in acht typen, waarvan de meeste hun eigen namen hebben en elk van hen wordt geproduceerd door een ander enzymatisch tekort. Sommigen zijn sterfelijk in zeer vroege levensfasen, terwijl anderen vergezeld gaan van spierzwakte en tekort tijdens het sporten.
Uitstekende voorbeelden
Enkele van de meest prominente glycogeengerelateerde ziekten zijn de volgende:
- Von Gierke's ziekte of glycogeenopslagziekte van type I, wordt geproduceerd door een 6-fosfatase glucosetekort in lever en nier.
Het wordt gekenmerkt door abnormale levergroei (hepatomegalie) als gevolg van de overdreven accumulatie van glycogeen en hypoglykemie, omdat de lever niet in staat is om glucose te leveren tot circulatie. Patiënten met deze aandoening hebben groeimotoraties.
- Pompe- of type II-ziekte is te wijten aan een α-tekort (1 → 4) -glucano 6-glycosyltransferas in de lever-, hart- en skeletspieren. Deze ziekte, zoals Andersen of Type IV, is dodelijk vóór de twee levensjaren.
- McArdle of Type V -ziekte heeft een spierfosforylase -tekort en gaat gepaard met spierzwakte, verminderde inspanningstolerantie, abnormale accumulatie van spierglycogeen en afwezigheid van lactaat tijdens het sporten tijdens inspanning.
Referenties
- Bhattacharya, K. (2015). Onderzoek en beheer van de leverglycogeenopslagziekten. Translationele kindergeneeskunde, 4(3), 240-248.
- Dagli, a., Sentner, c., & Weinstein, D. (2016). Glycogeenopslagziekte Type III. Gene Reviews, 1-16.
- Guyton, a., & Hall, J. (2006). Leerboek van medische fysiologie (11e ed.)). Elsevier Inc.
- Mathews, c., Van Holde, K., & Ahern, k. (2000). Biochemie (3e ed.)). San Francisco, Californië: Pearson.
- McKiernan, P. (2017). Pathobiologie van de wens van de leverglycogeenopslag. Curr Pathobiol Rep.
- Murray, r., Bender, D., Botham, k., Kennelly, p., Rodwell, V., & Weil, p. (2009). Harper's geïllustreerde biochemie (28e ed.)). McGraw-Hill Medical.
- Nelson, D. L., & Cox, m. M. (2009). Lehninger -principes van biochemie. Omega -edities (5e ed.)).
- Rawn, J. D. (1998). Biochemie. Burlington, Massachusetts: Neil Patterson Publishers.
- Tarnopolsky, m. NAAR. (2018). Myopathieën gerelateerd aan stoornissen van glycogeen metabolisme. Neurotherapeutica.
- « Argongeschiedenis, structuur, eigenschappen, gebruik
- Bijectieve functie Wat is, hoe wordt het gedaan, voorbeelden, oefeningen »