

Enolase structuur, werkingsmechanisme, functies
- 1479
- 34
- Alton D'Amore
De Versterken Het is het enzym dat wordt beschuldigd van het uitvoeren van de omzetting van D-2-fosfoglyceraat (2PGA) in fosfoenolpyruvaat (PEP) in glycolyse en omgekeerde reactie in gluconeogenese, twee metabole routes die deel uitmaken van de cellulaire energiemetabolisme.
De beslissing om deze reactie in een of andere richting te katalyseren, hangt af van de toegang die de cel tot glucose heeft. Dat wil zeggen van de behoeften van het aanpassen van uw metabolisme aan afbraak of synthese om energie te verkrijgen. Onmisbaar voor de realisatie van zijn vitale processen.
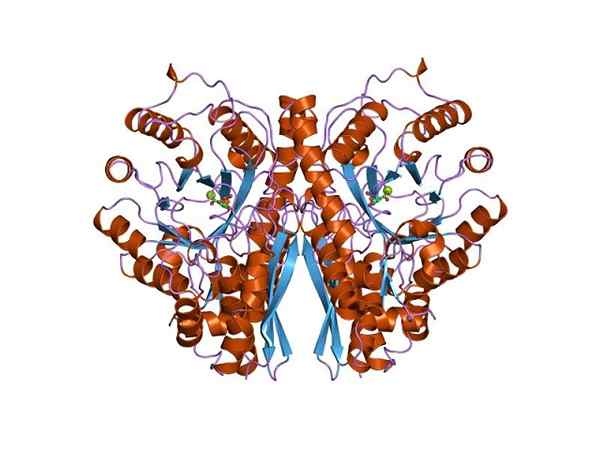
Drie -dimensionale structuur van enolasa. Door Jawahar Swaminathan en MSD -personeel van het European Bioinformatics Institute [Public Domain (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons. Aangezien beide metabole routes tot het centrum van de centrale metabole boom van levende wezens behoren, is het niet vreemd dat de aminozuursequentie van dit eiwit wordt bewaard in archeae, bacteriën en eukaryoten. En daarom presenteert dat vergelijkbare katalytische eigenschappen.
De locatie van enolase in de cel is beperkt tot cytosol, een compartiment waarin zowel glycolyse (ook wel glycolyse genoemd) als gluconeogenese in de meeste organismen plaatsvindt.
Het is echter ook gedetecteerd in andere celcompartimenten zoals plasmamembraan van veel kankercellen en cellen. Daar lijkt het betrokken te zijn bij de facilitering van celverspreidingsprocessen, een totaal andere functie dan zijn klassieke functie.
Enzymen die in staat zijn om meer dan één functie uit te voeren, zoals de enolase, staan bekend als Moonlighting -enzymen.
[TOC]
Structuur
De quaternaire structuur van de linus of niet in zijn liganden is bepaald in een groot aantal prokaryotische en eukaryotische individuen.
Elk monomeer presenteert twee domeinen: een klein amino-terminaal domein en een groter carboxyl-terminaal domein. Het N-terminale domein bestaat uit drie α en vier β-vellen. Terwijl de C-terminale bestaat uit acht β-vellen die afwisselen tussen hen die een β-vat vormen die is omgeven door acht α-propellers.
Bovendien worden twee vakbondslocaties voor divalente kationen gevonden in elk monomeer die "conformationele site" en "katalytische site" worden genoemd. De eerste is niet erg selectief en kan zich aansluiten bij een breed scala aan tweewaardige kationen in afwezigheid van substraat.
Kan u van dienst zijn: SPHINGOLIPIDS: wat zijn, kenmerken, functies, syntheseTerwijl de tweede bindt aan de ionen nadat het substraat zich heeft aangesloten bij het enzym. De unie van ionen op beide locaties is van vitaal belang voor de reactie om door te gaan.
Ten slotte is het belangrijk om te vermelden dat in de homodimeren de monomeren worden vergezeld door een parallelle oriëntatie te behouden. Daarom is de actieve site beperkt tot de centrale regio gevormd door genoemde Unie.
Slechts een van de twee monomeren neemt echter deel aan katalyse. Dit verklaart dat het vermogen van monomeren om de reactie uit te voeren onder experimentele omstandigheden.
Werkingsmechanisme
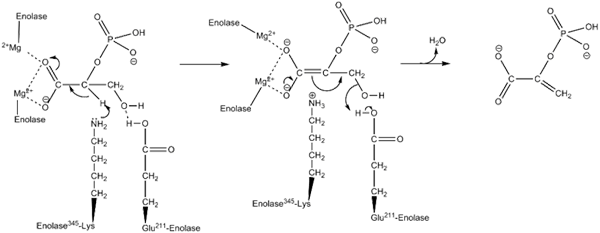 Werkingsmechanisme gebruikt door het enzym inolase. Door Kthompson08 in het Engels Wikipedia [Public Domain (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons.
Werkingsmechanisme gebruikt door het enzym inolase. Door Kthompson08 in het Engels Wikipedia [Public Domain (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons. Structurele studies, evenals die die hebben toegestaan om de kinetische en fysicochemische kenmerken van enolasa te bepalen, hebben ons in staat gesteld om hun werkingsmechanisme te begrijpen.
De manier waarop het enzym de katalyse van de reactie uitvoert, is behoorlijk interessant. Hoewel er een enkel substraat bij betrokken is, is een geordend sequentieel mechanisme dat is voorgesteld.
Dit begint met de vereniging van een ion van Mg2+ tot de conformationele site van een van de monomeren. Continu met de vereniging van het substraat op de actieve site gevolgd door de vereniging van een tweede ion naar de katalytische plaats en eindigt met de snelle afgifte van het product zodra de reactie is uitgevoerd. Op dit punt blijft de Mg2+ gekoppeld aan de conformationele site.
Langs dezelfde lijnen, om de realisatie van de reactie te bevorderen, het gemiddelde enzym in eerste instantie het genereren van een Carbanion -intermediair, waardoor een proton van koolstof 2 van de 2PGA wordt geëlimineerd. Dit doet dit dankzij de werking van een basisch aminozuurresidu.
Opeenvolgend vindt koolstof 3 hydroxylverwijdering plaats door de werking van een zuur residu uit het enzym. Op dit punt wordt de vereniging van beide koolstofatomen uitgevoerd door middel van een dubbele binding die PEP vormt. Op deze manier wordt de reactie geculmineerd.
Het kan u van dienst zijn: Monera Kingdom: Kenmerken, classificatie en voorbeeldenFunctie
Veel van de tot nu toe bestudeerde enzymen zijn in staat om een breed scala aan functies uit te voeren die niet gerelateerd zijn aan hun "klassieke functie" in verschillende cellulaire compartimenten. Deze enzymen worden "maanlicht" enzymen genoemd.
In deze zin kan enolase worden beschouwd als een enzym maanlicht, omdat talloze functies tegen hun klassieke functie zijn toegeschreven tot op heden, zowel in bacteriën als in eukaryoten.
Sommige van deze functies zijn de volgende:
- Deelnemen aan het behoud van celvorm en aan vesiculair verkeer bij interactie met cytoskeleteiwitten.
- In de kern van zoogdiercellen werkt als een transcriptiefactor die de expressie reguleert van genen geassocieerd met celproliferatie. Samenwerken bij het behoud van de stabiliteit van de RNM in de afbraak in bacteriën.
- In ziekteverwekkers, zoals Streptococcus pneumoniae En Trypanosoma cruzi, Het lijkt erop dat het fungeert als een belangrijke virulentiefactor.
- Er is ook gebleken dat in Streptococcus pyogenes, Enolase wordt uitgescheiden aan het extracellulaire medium dat weefselafbraak en de ontwijking van het gastheiligssysteem vergemakkelijkt.
- Op het oppervlak van tumorcellen wordt uitgedrukt door het verbeteren van metastase.
Enolase en zijn relatie met celverwijderingsmechanismen
Talrijke pathogenen, evenals tumorcellen, brengen in hun membraan uit of scheiden de extracellulaire middelgrote proteasen uit die in staat zijn eiwiteiwitten af te breken.
Met deze capaciteit kunnen deze cellen door de weefsels breken en zich snel verspreiden over het hele gastheerorganisme. Dus de voorkeur geven aan de ontwijking van het immuunsysteem en daarom de oprichting van de infectie.
Zelfs wanneer enolasa protease -activiteit mist, neemt deel aan het proces van het verspreiden van vele pathogenen in zijn gastheer en tumorcellen tijdens doelstastase.
Dit bereikt het dankzij het feit dat het tot expressie wordt gebracht op het oppervlak van deze cellen die functioneert als een plasminogeenreceptor. De laatste is het zimogeen van een serin-prototease die bekend staat als plasminaat dat deel uitmaakt van het fibrinolytische systeem en werkt afbraak van extracellulaire matrixeiwitten.
Daarom is de op het oppervlak tot expressie gebrachte enolase een strategie die deze cellen hebben verkregen om de infectie vast te stellen en zich succesvol te verspreiden.
Het kan u van dienst zijn: Sympatric Speciation: Concept, Kenmerken en voorbeeldenDeze strategie bestaat uit twee processen:
- De ontwijking van het immuunsysteem van de gastheer. Omdat, wanneer deze cellen van een gastheer -eiwit worden gecoat, ze worden genegeerd door cellen van het immuunsysteem die niet -eigenaar pathogenen herkennen die geassocieerd zijn met ziekteverwekkers.
- Verspreiding na de actie van plasminogeen in plasminaat. Waarvan de deelname aan de afbraak van extracellulaire matrixeiwitten en vervolgens de snelle en effectieve verspreiding vergemakkelijkt.
Referenties
- Avilan L, Gualdron-Lopez M, Quiñones W, González-González L, Hannaert V, Michels PAA, Concepción Jl. Enolase: een belangrijke speler in het metabolisme en een waarschijnlijke virulentiefactor van trypanosomatid parasieten-perspectieven voor het gebruik ervan als therapeutisch doelwit. Enzymonderzoek. 2011 Vol. Artikel ID932549, 14 pagina's.
- Bhowmick I, Kumar N, Sharma S, Coppens I, Jarori GK, Plasmodium falciparum enosoase: stadium-specifieke expressie en subcellulaire lokalisatie. Malaria -dagboek. 2009; 8 (1). Artikel 179.
- Dag I, Peshavaria M, Quinn GB, een differentiële moleculaire klok in Enoase isoproteïne -evolutie. Journal of Molecular Evolution. 1993; 36 (6): 599-601.
- van de Torre-Scuder E, Manzano-Román R, Pérez-Sánchez R, Siles-Lucas M, Oleagaa a. Klonering en karakterisering van plasminogeen -bindend oppervlakte -geocieerde enolaas Schistosoma Bovis. Veterinaire parasitologie. 2010; 173: 73-84.
- Dinovo EC, Boyer PD. Isotopische sondes van het enolase -reactiemechanisme. Initiële en equiquibium isotoopwisselingsrente: primaire en secandaire isotoopeffecten. J Biol Chem. 1971; 246 (14): 4586-4593.
- Kaberdin VR, Lin-Chao S, het ontrafelen van nieuwe rollen voor kleine componenten van de E. Coli RNA degradueren. RNA -biologie. 2009; 6 (4): 402-405.
- Keller A, Peltzer J, Carpentier G. Interacties van enolas. Biochimica et Biophysica Acta. 2007; 1770 (6): 919-926.
- Lung J, Liu KJ, Chang JY, Leu SJ, Shih NY. MBP-1 wordt effeciënt gecodeerd door een alternatief transcript van het Enri1-gen buttranslationeel gereguleerd door proteasoom-afhankelijke eiwitverschuiving. Febs Journal. 2010; 277 (20): 4308-4321.
- Pancholi V. Multifunctionele α-enolase: zijn rol in ziekten. Cellulaire en moleculaire levenswetenschappen. 2001; 58 (7): 902-920.
- Poyner RR, Cleland WW, Reed GH. Rol van metaalins in katalyse door enolase. Een geordend kinetisch mechanisme voor een enkel substraat -enzym. Biochemie. 2001; 40: 9008-8017.
- Segovia-Gamboa NC, Chávez-Munguía B, Medina-Flores A, Entamoeba komt binnen, Encystation -proces en enolase. Experimentele parasitologie. 2010; 125 (2): 63-69.
- Tanaka M, Sugisaki K, Nakashima K, schakelen in niveaus van vertaalbare mRNA's voor Enoase -isozymen tijdens de ontwikkeling van Chickn -skeletspier. Biochemische en biofysische onderzoekscommunicatie. 1985; 133 (3): 868-872.