

Vertaling van het DNA -proces in eukaryotas en prokaryoten
- 4818
- 1226
- Kurt Aufderhar Jr.
De DNA -vertaling Het is het proces waarmee de informatie in de boodschappers die tijdens transcript zijn geproduceerd, opgenomen.
Vanuit cellulair perspectief is de expressie van een gen een relatief complexe materie die in twee stappen optreedt: transcriptie en vertaling.
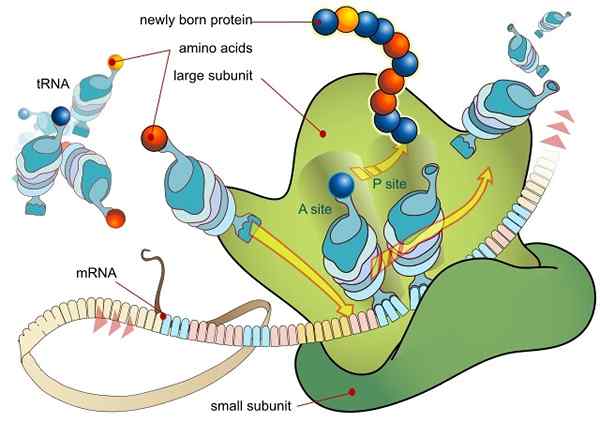
 RNA -vertaling gemedieerd door een ribosoom (Bron: Ladyofhats / Public Domain, via Wikimedia Commons)
RNA -vertaling gemedieerd door een ribosoom (Bron: Ladyofhats / Public Domain, via Wikimedia Commons) Alle genen die tot expressie worden gebracht (of het nu al dan niet coderen voor peptidesequenties, dat wil zeggen eiwitten), doen dit aanvankelijk door de informatie over te dragen in zijn DNA -sequentie naar een messenger mer molecula (RNAM) via een proces dat transcriptie wordt genoemd.
Het transcript wordt bereikt door speciale enzymen die bekend staan als RNA-polymerasen, die een van de complementaire strengen van het DNA van het gen gebruiken als een vorm voor de synthese van een "pre-arnm" molecuul, dat vervolgens wordt vervolgd om een volwassen volwassen te vormen.
Voor genen die coderen voor eiwitten, wordt de informatie in volwassen RNM "gelezen" en vertaald in aminozuren volgens de genetische code, die aangeeft welk codon- of nucleotide -triplet overeenkomt met elk specifiek aminozuur.
De specificatie van de aminozuursequentie van een eiwit hangt daarom af van de initiële sequentie van stikstofbasen in het DNA dat overeenkomt met het gen en vervolgens in het RNA dat genoemde informatie van de kern naar het cytosol transporteert (in eukaryotische cellen); proces dat ook wordt gedefinieerd als de synthese van eiwit geleid door RNM.
Gezien het feit dat er 64 mogelijke combinaties zijn van de 4 stikstofbases die DNA en RNA vormen en slechts 20 aminozuren, kan een aminozuur worden gecodeerd door verschillende drieling (codons), dus er wordt gezegd dat de genetische code is " gedegenereerd "(behalve de aminozuurmetionine, die wordt gecodeerd door een uniek aug -codon).
[TOC]
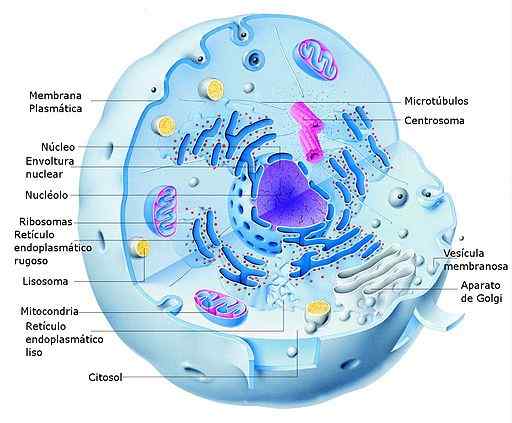
Eukaryota-vertaling (stappenprocessen)
Diagram van een eukaryotcel van dieren en zijn delen (bron: Alejandro Porto [CC0] via Wikimedia Commons) In eukaryotische cellen vindt de transcriptie plaats in kern en de vertaling in het cytosol, zodat de RNM's die worden gevormd tijdens het eerste proces ook een functie vervullen in het transport van de informatie van de kern naar de cytosol, waar de biosynthetische machines (ribosomen (ribosomen worden )).
Het is belangrijk om te vermelden dat de compartimentering van transcriptie en vertaling in eukaryoten waar is voor de kern, maar het is niet hetzelfde voor organellen met hun eigen genoom zoals chloroplasten en mitochondria, die meer vergelijkbare systemen hebben als die van lekkaryotische organismen.
Eukaryotische cellen hebben ook cytosolische ribosomen bevestigd aan de membranen van het endoplasmatische reticulum (ruw endoplasmatisch reticulum), waarbij de translatie van de eiwitten die zijn bedoeld om in celmembranen te worden ingevoegd of die de post -translationele verwerking vereisen die optreden voorkomt.
- RNM -verwerking voorafgaand aan hun vertaling
RNM's worden aan hun uiteinden gewijzigd als ze worden getranscribeerd:
- Wanneer het 5 'uiteinde van de RNM tijdens de transcriptie uit het oppervlak van de RNA-polymerase II komt, wordt dit onmiddellijk "aangevallen" door een groep enzymen die een "kap" die bestaat uit 7-methylguanilado en die is verbonden met de terminal Nucleotide van de RNM via een 5 'triffosfaatverbinding, 5'.
Kan u van dienst zijn: codon- Het 3 'uiteinde van de RNM lijdt aan een "clivaje" door een endonuclease, dat een gratis hydroxylgroep 3' genereert die zich aansluit bij een "ristra" of "staart" van adenine -afval (van 100 tot 250) die er een worden toegevoegd. Tijd voor een enzym Poly (A) Polymerase.
De "kap 5 '" en de "staart Poly A ”Ze vervullen functies in de bescherming van RNM -moleculen tegen afbraak en werken ze bovendien in het transport van volwassen transcripten naar de cytosol en in de initiatie en beëindiging van de vertaling, respectievelijk.
COrte en empalme
Na de transcriptie passeren de "primaire" RNM's met hun twee gemodificeerde uitersten, nog steeds aanwezig in de kern, een "snij- en splitsing" -proces waarmee intronics -sequenties in het algemeen worden geëlimineerd en de resulterende exons worden verbonden (post -regionale verwerking), Met wat de volwassen transcripties die de kern verlaten en het cytosol bereiken, worden verkregen.
De snit en splitsing wordt uitgevoerd door een riboproteisch complex genaamd de Espicleosoma (Anglicisme van Spliceosome), gevormd door vijf kleine ribonucleoproteïnen en RNA -moleculen, die in staat zijn om de gebieden te "herkennen" die moeten worden geëlimineerd uit het primaire transcript.
In veel eukaryoten is er een fenomeen dat bekend staat als "snijden en alternatief gewricht", wat betekent dat verschillende soorten post -registratie -modificaties verschillende of isoenzyme -eiwitten kunnen veroorzaken die van elkaar verschillen in sommige aspecten van hun sequenties.
- De ribosomen
Wanneer volwassen transcripten de kern verlaten en worden getransporteerd voor translatie in cytosol, worden deze verwerkt door het translationele complex dat bekend staat als ribosoom, dat bestaat uit een eiwitcomplex geassocieerd met RNA -moleculen.
De ribosomen zijn samengesteld uit twee subeenheden, een "groot" en een andere "klein", die vrijelijk worden gedissocieerd in het cytosol en zich aansluiten bij het mRNA -molecuul dat zich vertaalt.
De vereniging tussen de ribosomen en het mRNA hangt af van gespecialiseerde RNA -moleculen die worden geassocieerd met ribosomale eiwitten (ribosomaal RNA of RNA en overdracht of arnt RNA), die elk specifieke functies uitoefenen.
De arnt zijn moleculaire "adapters", omdat ze door een van hun uiteinden elk codon of triplet kunnen "lezen" in het volwassen RNA (door complementariteit van bases) en door de andere kunnen ze zich aansluiten bij het aminozuur gecodeerd door het codon "gelezen".
RNR -moleculen zijn daarentegen verantwoordelijk voor het versnellen (katalyseren) van het bindingsproces van elk aminozuur in de ontluikende peptideketen.
Een eukaryotische volwassen RNM kan door vele ribosomen worden "gelezen", zo vaak als de cel dit aangeeft. Met andere woorden, dezelfde RNM kan leiden tot veel exemplaren van hetzelfde eiwit.
Codon en leeskader starten
Wanneer een volwassen RNM wordt benaderd door de ribosomale subeenheden, "scan" het riboproteïne complex de volgorde van genoemde molecuul totdat een startcodon zich bevindt, wat altijd augustus is en impliceert de introductie van een methionineresidu.
Het kan je van dienst zijn: monoploidy: hoe het gebeurt, organismen, frequentie en nutHet aug -codon definieert het leeskader voor elk gen en definieert bovendien het eerste aminozuur van alle in de natuur vertaalde eiwitten (dit aminozuur wordt vaak post -translationeel geëlimineerd).
Beëindigingscodons
Drie andere codons zijn geïdentificeerd als die welke vertalende beëindiging veroorzaken: UAA, UAG en UGA.
Die mutaties die een verandering van stikstofbases in het triplet impliceren die codeert voor een aminozuur en die resulteren in beëindigingscodons staan bekend als betekenisloze mutaties, omdat ze voortijdige detentie van het syntheseproces veroorzaken, dat kortere kortere kortere eiwitten vormt.
Niet -vertaalde regio's
Nabij het 5 'einde van de volwassen RNM -moleculen zijn er regio's die niet worden vertaald (UTR, uit het Engels Niet -vertaalde regio), ook wel 'leider' -sequenties genoemd, die zich tussen de eerste nucleotide en het begin van de vertaling bevinden (aug).
Deze UTR -regio's die niet worden vertaald, hebben specifieke locaties voor vereniging met ribosomen en mensen, bijvoorbeeld, ze hebben een geschatte lengte van 170 nucleotiden, waaronder regulerende gebieden, eiwitbindingsplaatsen die werken bij de regulatie van de vertaling, enz.
- Start van vertaling
De vertaling, evenals de transcriptie bestaat uit 3 fasen: een van initiatie, een andere van verlenging en uiteindelijk een van beëindiging.
Initiatie
Het bestaat uit de assemblage van het translatiecomplex op de RNM, die de vereniging verdient van drie eiwitten die bekend staan als initiatiefactoren (als, van het Engels Initiatiefactor) IF1, IF2 en IF3 naar de kleine subeenheid van het ribosoom.
Het "pre -initiatie" -complex gevormd door de initiatiefactoren en de kleine ribosomale subeenheid worden op hun beurt verbonden met een arnt dat "een metie residu" laadt en deze set moleculen bindt aan de RNAM, nabij het startcodon aug.
Deze gebeurtenissen leiden tot de RNM -unie met de grote ribosomale subeenheid, die leidt tot de afgifte van initiatiefactoren. De grote subeenheid van het ribosoom heeft 3 vakbondsplaatsen voor ARNT -moleculen: site A (aminozuur), site P (polypeptide) en site E (output).
Site A sluit zich aan bij de aminoacil-arnt anticod, die complementair is aan dat van het vertaalde mRNA; De P -site is waar het aminozuur wordt overgebracht van de arnt naar het opkomende peptide en de S -site is waar het in arnt "leeg" is voordat het wordt vrijgegeven aan de cytosol na het leveren van het aminozuur.
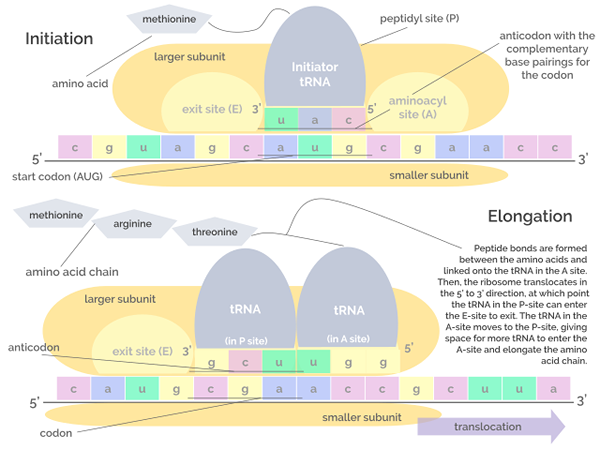 Grafische weergave van de initiatie- en verlengingsfasen van de vertaling (Bron: Jordan Nguyen/CC BY-S (https: // CreativeCommons.Org/licenties/by-sa/4.0) Via Wikimedia Commons)
Grafische weergave van de initiatie- en verlengingsfasen van de vertaling (Bron: Jordan Nguyen/CC BY-S (https: // CreativeCommons.Org/licenties/by-sa/4.0) Via Wikimedia Commons) Verlenging
Deze fase bestaat uit de "beweging" van ribosoom langs het mRNA -molecuul en de translatie van elk codon dat "lezen" wat de groei of verlenging van de polypeptideketen in geboorte impliceert.
Dit proces vereist een factor die bekend staat als de verlengingsfactor van GTP in de vorm van GTP, die de translocatie van verlengingsfactoren langs het RNM -molecuul stimuleert terwijl het vertaalt.
Kan u van dienst zijn: Okazaki -fragmentenDe transferase peptidilactiviteit van ribosomale RNA's maakt de vorming van peptidebindingen mogelijk tussen opeenvolgende aminozuren die aan de keten worden toegevoegd.
Beëindiging
De vertaling eindigt wanneer het ribosoom aan een van de beëindigingscodons voldoet, omdat de ARN's deze codons niet herkennen (ze coderen geen aminozuren). Eiwitten die bekend staan als afgiftefactoren worden ook verbonden, die het onthechting van het ribosoma mr mogelijk maken.
Procary vertaling (passes-processen)
In prokaryoten, zoals in eukaryotische cellen, worden de ribosomen die op lading van eiwitsynthese worden aangetroffen in cytosol (wat ook waar is voor transcriptionele machines), een feit dat de snelle toename van de cytosolische concentratie van een eiwit mogelijk maakt wanneer de expressie van de genen dat Codeert dat het toeneemt.
Hoewel het geen extreem veel voorkomend proces is in deze organismen, kan de primaire RNM die tijdens transcriptie wordt geproduceerd na -registratie -rijping door "snijden en splitsen" lijden ". De meest voorkomende is echter om ribosomen te observeren die zijn bevestigd aan de primaire getranscribeerde die het vertalen op hetzelfde moment dat het wordt getranscribeerd uit de overeenkomstige DNA -sequentie.
Gezien het bovenstaande begint de vertaling in veel prokaryoten met het einde 5 ', omdat het 3' uiteinde van het mRNA blijft verbonden met het schimmel -DNA (en komt samen met de transcriptie voor).
Niet -vertaalde regio's
Prokaryotische cellen produceren ook RNM met niet-vertaalde gebieden die bekend staan als de "Shine-Dalgarno Box" en waarvan de consensussequentie Aggagg is. Zoals duidelijk is, zijn de UTR -gebieden van bacteriën aanzienlijk korter dan die van eukaryotische cellen, hoewel ze vergelijkbare functies uitoefenen tijdens vertaling.
Proces
In bacteriën en andere prokaryotische organismen is het translatieproces vrij gelijkaardig aan dat van eukaryotische cellen. Het bestaat ook uit drie fasen: initiatie, verlenging en beëindiging, die afhankelijk zijn van specifieke prokaryotische factoren, verschillend van die welke door eukaryoten worden gebruikt.
Verlenging hangt bijvoorbeeld af van verlengingsfactoren die bekend staan als EF-TU en EF-TS, in plaats van de eukaryotische factor G.
Referenties
- Alberts, B., Johnson, a., Lewis, J., Raff, m., Roberts, K., & Walter, p. (2007). Biologie van de celmoleculair. Slingerwetenschap. New York, 1392.
- Klei, s. & Brown, W. (2008) Vertaling: DNA naar mRNA naar eiwit. Nature Education 1 (1): 101.
- Griffiths, een. J., Wessler, s. R., Lewontin, r. C., Gelbart, W. M., Suzuki, D. T., & Miller, J. H. (2005). Een inleiding tot genetische analyse. Macmillan.
- Lodish, h., Berk, een., Kaiser, c. NAAR., Krieger, m., Scott, m. P., Bretscher, een.,… & Matsudaira, p. (2008). Moleculaire celbiologie. Macmillan.
- Nelson, D. L., Lehninger, een. L., & Cox, m. M. (2008). Lehninger -principes van biochemie. Macmillan.
- Rosenberg, l. EN., & Rosenberg, D. D. (2012). Menselijke genen en genomen: wetenschap. Health, Society, 317-338.
- « Sorbus aria -kenmerken, habitat, eigenschappen, cultuur
- Boletus Aereus -functies, habitat, identificatie, recepten »