

Bot Fabric -kenmerken, structuur, vorming en groei
- 2287
- 164
- Dr. Rickey Hudson
Hij geweven bot is degene die de botten samenstelt. Het bot, samen met email en dentine, zijn de moeilijkste stoffen in het lichaam van de dieren. De botten vormen de structuren die de vitale organen beschermen: de hersenen worden beschermd door de schedel, het ruggenmerg door de wervelkolom en het hart en de longen zijn door de ribbenkast.
De botten dienen ook als "hefbomen" voor de spieren die erin worden ingevoegd, waardoor de kracht die deze spieren genereren tijdens de uitvoering van de bewegingen vermenigvuldigen. De stijfheid van het bot maakt de voortbeweging en ondersteuning van de belastingen tegen de zwaartekracht mogelijk.
Botweefselcellen (Bron: OpenStax College [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons) Het bot is een dynamisch levende stof die constant verandert en deze veranderingen worden gestimuleerd door de druk en spanningen waaraan dit weefsel wordt onderworpen. Druk stimuleert bijvoorbeeld resorptie (vernietiging) en spanning kunnen nieuwe botvorming stimuleren.
De botten vormen de belangrijkste afzetting van calcium en fosfor van organisme: bijna 99% van het totale calcium van het menselijk lichaam wordt opgeslagen in het botweefsel. Totale botmassa varieert gedurende de hele levensduur van een dier. Tijdens de groeifase overschrijdt botvorming de resorptie (vernietiging) en het skelet groeit en ontwikkelt zich.
Aanvankelijk neemt de lengte toe en vervolgens zijn dikte, waardoor het maximaal wordt bereikt tussen de 20 en 30 jaar bij mensen. In de volwassene (tot ongeveer 50 jaar) is er een evenwicht tussen vorming en botresorptie.
Dit saldo wordt gegeven door een vervangingsproces dat bekend staat als "botremodellering" en dat per jaar treft van 10% tot 12% van de totale botmassa. Vervolgens begint een degeneratief proces waarbij de resorptie de vorming overschrijdt en de botmassa langzaam afneemt.
[TOC]
Kenmerken en structuur
Het bot heeft een centrale holte die kernholte wordt genoemd, die het beenmerg herbergt, een hematopoietisch weefsel, dat wil zeggen een bloedcel dat weefsel vormt. Deze structuren worden gedekt door periosteum, met uitzondering van de gebieden die overeenkomen met de synoviale gewrichten.
Het periosteium heeft een buitenste laag vezelachtig dicht bindweefsel en een binnenste laag met osteogene cellen, die botvormende cellen of osteoprogencellen zijn.
Het centrale deel van het bot wordt gestoffeerd door een dunne en gespecialiseerde bindweefselcelmonolay. Endostio heeft osteoprogen en osteoblasten -cellen. Het aldus gestoffeerde bot heeft zijn cellen geïntegreerd in een matrix [F1] [F2] extracellulaire verkalkte.
Osteoprogenitorcellen verschillen in osteoblasten en zijn belast met het afscheiden van de botmatrix. Wanneer omringd door matrix, worden deze cellen geïnactiveerd en is de naam van osteocyten geïnactiveerd.
De ruimtes bezet door osteocyten in de matrix worden lagunes genoemd.
90% van de organische matrix wordt gevormd door type I collageenvezels, een structureel eiwit dat ook aanwezig is in pezen en huid, en de rest is een homogene jelly -substantie die fundamentele substantie wordt genoemd.
Compact bot en sponsachtig bot
De collageenvezels van de matrix zijn gerangschikt in grote balken en in het compacte bot vormen deze vezels concentrische lagen rond de kanalen waardoor de bloedvaten en zenuwvezels (Havers Canals) run (havers). Deze lagen vormen cilinders bekend als "osteones".
Elke osteone wordt afgebakend door een cementatielijn gevormd door fundamentele substantie verkalkt met weinig collageenvezels en voedt de vaten die zich in de Hovers -kanalen bevinden.
In het sponsachtige bot worden grote plaques of spicules gevormd en worden de cellen gevoed door diffusie van de extracellulaire vloeistof van het bot naar de trabeculae.
De anorganische componenten van de matrix vormen ongeveer 65% van het droge gewicht van het bot en worden voornamelijk gevormd door calcium en fosfor, naast sommige elementen zoals natrium, kalium, magnesium, citraat en bicarbonaat, onder anderen.
Kan u van dienst zijn: Merychippus: kenmerken, reproductie, voeding, taxonomieCalcium en fosfor vormen hydroxyapatietkristallen [CA10 (PO4) 6 (OH) 2]. Calciumfosfaat wordt ook gevonden in amorfe vorm.
Hydroxyapatietkristallen zijn op orde gerangschikt.
Botvorming en groei
De schedelbotten worden gevormd door een proces dat bekend staat als "Intramembranous Ossification". In plaats daarvan worden lange botten eerst gemodelleerd in kraakbeen en vervolgens omgezet in bot door ossificatie, die begint in de diafyse van het bot en "endochondrale ossificatie" wordt genoemd.
De meeste platte botten ontwikkelen en groeien door intramembranous botvorming en ossificatie. Dit proces vindt plaats in het zeer gevasculariseerde mesenchymale weefsel, waarbij mesenchymale cellen verschillen in osteoblasten die beginnen met het produceren van botmatrix.
Dit is hoe een netwerk van spicules en trabeculae wordt gevormd, waarvan de oppervlakken worden bevolkt door osteoblasten. Deze initiële osteogenese -regio's worden primair ossificatiecentrum genoemd. Dit vormt het primaire bot met willekeurig georiënteerde collageenvezels.
Dan worden de verkalking en de osteoblasten die in de matrix gevangen zitten, osteocyten, waarvan de uitbreidingen aanleiding geven tot kanalen. Omdat trabeculaire netwerken zich vormen als een spons, geeft vasculair bindweefsel aanleiding tot het beenmerg.
De toevoeging van perifere trabeculae verhoogt de botgrootte. In het occipitale bot (een schedelbot in de achterste zone) zijn er verschillende ossificatiecentra die samenvoegen om een enkel bot te vormen.
Bij pasgeborenen zijn sanitair tussen de voor- en pariëtale botten ossificatiezones die nog niet zijn samengevoegd.
Compacte botvorming
De MesenQuimatous Tissue -gebieden die zonder verkalking in interne en externe delen blijven, vormen het periosteum en endostio. De sponsachtige botgebieden onmiddellijk naar het periosteum en de Duramadre worden compact bot en vormen de interne en externe tabel van het platte bot.
Tijdens de groei, in lange botten, worden gebieden die gespecialiseerd zijn in epifyses gescheiden van de diafyse door een zeer actieve kraakbeenplaat genaamd epifysische plaat.
De botlengte neemt toe in de mate dat deze plaat nieuw bot afzet aan elk uiteinde van de diafyse. De grootte van de epiphysplaat is evenredig met de groeisnelheid en wordt beïnvloed door verschillende hormonen.
Regulatie
Onder de hormonen die deze plaat moduleren, is het groeifatgebied (GH) dat wordt vrijgegeven door de vorige hypofyse-groei vergelijkbaar met type I insuline (IGF-I) geproduceerd door de lever.
Hoewel de mitotische activiteitssnelheid in de proliferatiezone vergelijkbaar is met botresorptiesnelheid van het gebied, blijft de grootte van de epifysplaat constant en blijft het bot groeien.
Na 20 jaar neemt de mitotische activiteit af en bereikt de ossificatiezone de kraakbeenzone, waarbij de kernholten van diafyse en epifyses worden verbonden.
De longitudinale groei van het bot eindigt wanneer de epiphysele sluiting optreedt, dat wil zeggen wanneer de diafyse wordt verbonden met epifyses. De epiphysenale sluiting volgt een geordende tijdelijke volgorde die eindigt met de laatste sluiting na de puberteit.
De breedtegroei van het lange bot wordt geproduceerd door apostale groei, wat het product is van de differentiatie van de osteoprogenitorcellen van de binnenste laag van het periosteum in osteoblasten die botmatrix afscheiden op de subperiostische gebieden van de diafyse.
Bot -remodellering
Gedurende het leven van een mens wordt het bot voortdurend vervangen door de vorming- en resorptieprocessen, dat wil zeggen van vernietiging van het oude bot en de nieuwe botvorming.
Het kan u dienen: Sinaloa Flora en Fauna: Meer gewone dieren en plantenBij zuigelingen lijdt calcium een 100% jaarlijkse vervanging, terwijl bij volwassenen het slechts 18% per jaar is. Deze resorptie en vorming of vervangingsprocessen worden botremodellering genoemd.
De remodellering begint met de werking van de osteoclasten die het bot vernietigen en enkele kloven achterlaten die vervolgens worden binnengevallen door de osteoblasten. Deze osteoblasten scheiden de matrix uit die vervolgens zal worden verzorven en geeft aanleiding tot het nieuwe bot. Deze cyclus vereist gemiddeld meer dan 100 dagen.
Op een bepaald tijdstip is min of meer 5% van de gehele botmassa van het skelet tijdens het remodelleren. Dit impliceert de deelname van ongeveer twee miljoen remodellerende eenheden.
Verschillen in de remodellering van het compacte en sponsachtige bot
Het jaarlijkse compacte bottenremodelleringspercentage is 4 % en sponsachtige bot is 20 %.
Het verschil tussen de remodelleringssnelheden van de twee soorten bot is waarschijnlijk te wijten aan het sponsachtige bot in contact met het beenmerg en wordt direct beïnvloed door de cellen met paracriene activiteit van genoemde medulla.
De osteoprogencellen van de compacte botten worden integendeel gevonden in de Bearsiaanse kanalen en in de binnenste lagen van het periosteum, ver van de cellen van het beenmerg en hangen af voor het begin van de remodellering, van de hormonen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen die aankomen, door bloed.
Velen zijn de hormonale en eiwitfactoren die betrokken zijn bij de activiteit van osteoblasten en osteoclasten bij het remodelleren.
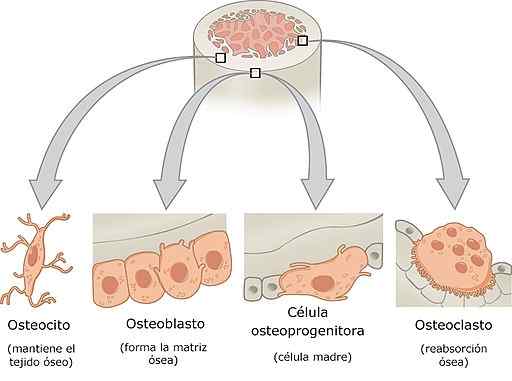
Bottencellen
-Soorten botcellen en hun kenmerken
Botcellen zijn osteoprogenitorcellen, osteoblasten, osteocyten en osteoclasten. Elk van deze cellen heeft specifieke functies in de botfysiologie en hebben zeer gedifferentieerde histologische kenmerken.
Osteoblasten, osteocyten en osteoclasten vormen samen de botmodeleenheid.
Osteoprogen of osteogene cellen
Deze cellen worden gevonden in de binnenste laag van het periosteum en in de endostio. Ze komen voort uit de embryonale mesenchym en kunnen aanleiding geven tot differentiatie, tot osteoblasten. Onder bepaalde stressomstandigheden kunnen ze ook differentiëren in chondrogene cellen.
Het zijn spindelvormige cellen met een ovale kern, schaars cytoplasma, met weinig ruw endoplasmatisch reticulum (RER) en een slecht ontwikkeld Golgi -apparaat. Ze hebben overvloedige ribosomen en zijn zeer actief tijdens de botgroeiperiode.
De osteoblasten
Osteoclasten zijn cellen afgeleid van osteogene cellen. Ze zijn verantwoordelijk voor het synthetiseren van de organische matrix van het bot, dat wil zeggen collageen, proteoglycans en glycoproteïnen. Ze zijn gerangschikt in lagen bovenop het botoppervlak.
De kern bevindt zich aan de andere kant van het secretoire gedeelte rijk aan blaasjes. Ze hebben een overvloedige RER en een goed ontwikkeld Golgi -apparaat. Ze hebben korte projecties of extensies die contact maken met andere aangrenzende osteoblasten. Andere lange extensies verbinden ze met osteocyten.
Omdat de osteoblasten ouder worden.
Hoewel het grootste deel van de botmatrix wordt verkalkt, is er rond elke osteoblast en zelfs elke osteocyt een dunne laag niet -gecalcificeerde botmatrix die osteoïde wordt genoemd en die deze cellen scheidt van de verkalkte matrix.
In het celmembraan van osteoblasten zijn er verschillende soorten receptoren. Van deze receptoren is de belangrijkste de ontvanger voor het parathyroïde hormoon (PTH), dat de secretie stimuleert van een stimulerende osteoclastfactor die botresorptie bevordert.
Osteoblasten kunnen ook enzymen scheiden die in staat zijn osteoïde te verwijderen en dus osteoclasten met het verkalkte botoppervlak te plaatsen om de resorptie te starten.
Osteocyten
Dit zijn cellen afgeleid van inactieve osteoblasten en worden volwassen botcellen genoemd. Ze zijn ondergebracht in de bovengenoemde lippen van de verkalkte botmatrix. Er zijn er tussen de 20.000 tot 30.000 osteocyten per kubieke millimeter bot.
Kan u dienen: fosfatidylcholine: synthese, structuur, functies, eigenschappenVan de lagunes stralen de osteocyten cytoplasmatische extensies uit die ze met elkaar verenigen, waardoor interstitium -vakbonden worden gevormd waarvoor ionen en kleine moleculen kunnen worden uitgewisseld tussen de cellen tussen de cellen.
Osteocyten zijn afgeplatte cellen, met platte en weinig cytoplasmatische organellen kernen. Ze zijn in staat om stoffen af te scheiden op mechanische stimuli die botspanning veroorzaken (transductiemechano).
De ruimte die de osteocyten in de lagunes omringt, wordt de periosteocytische ruimte genoemd en zit vol met extracellulaire vloeistof in de niet -gecalcificeerde matrix. Naar schatting is het oppervlak van de periosteocytenwanden ongeveer 5000 m2 en dat een volume van ongeveer 1,3 liter extracellulaire vloeistof herbergt.
Deze vloeistof wordt blootgesteld aan ongeveer 20 g uitwisselbaar calcium dat kan worden geresorbeerd in de bloedsomloopstroom uit de wanden van deze ruimtes, wat bijdraagt aan het behoud van de bloedfiguren van calcium.
Osteoclasten
Deze cellen zijn afkomstig van dezelfde voorlopercellen als weefselmacrofagen en circulerende monocyten; Deze worden gevonden in het beenmerg en zijn de voorlopercellen van granulocyten en macrofagen (GM-CFU).
De myitose van deze voorlopercellen wordt gestimuleerd door de stimulerende factoren van macrofaagkolonies en in aanwezigheid van botten, deze voorlopersfusie en vormen multinucleaire cellen.
Een osteoclast is een grote, multinucleaire en mobiele cel. Het meet een diameter van ongeveer 150 urn en kan tot 50 kernen hebben. Het heeft een basaal gebied waar de kernen en de organellen zich bevinden, een borstelrand in contact met het verkalkte bot, heldere perifere gebieden naar de borstelrand en een vesiculair gebied.
De belangrijkste functie van deze cellen is die van botresorptie. Zodra ze hun functie uitoefenen, lijden ze op apoptose (geprogrammeerde celdood) en sterven. Om het botresorptieproces te initiëren, houdt de osteoclast zich aan het bot door uitgebreide eiwitten.
Vervolgens bewegen protonen bommen die H+afhankelijk zijn van Atasas, van de endosomen naar de binnenkant van het membraan op een borstelrand en verzuren het medium totdat de pH ongeveer ongeveer 4 valt.
Hydroxyapatiet lost op aan dergelijke pH en collageenvezels worden afgebroken door zure proteasen die ook door deze cellen worden uitgescheiden. De eindproducten van de digestie van hydroxyapatiet en collageen zijn endocyed in de osteoclast en worden vervolgens vrijgegeven aan de interstitiële vloeistof om vervolgens te worden geëlimineerd door de urine.
Botweefseltypen (bottypen)
Zoals je misschien in de tekst hebt opgemerkt, zijn er twee soorten botweefsel, namelijk: het compacte of corticale bot en het trabeculaire of sponsachtige bot.
De eerste vormt 80% van de totale botmassa en wordt gevonden in de diafyse van de lange botten, die de buisvormige delen zijn die zijn gerangschikt tussen de twee uiteinden (epifyses) van deze botten.
Het tweede type bot is typerend voor de botten van het axiale skelet, zoals de wervels, de botten van de schedel en het bekken en de ribben. Het bevindt zich ook in het midden van lange botten. Het vormt 20% van de totale botmassa en is van vitaal belang voor de regulatie van calciummetabolisme.
Referenties
- Berne, r., & Levy, m. (1990). Fysiologie. Mosby; Internationale editie.
- Di fiore, m. (1976). Normale histologie -atlas (2e ed.)). Buenos Aires, Argentinië: de redactie Athenaeum.
- Doubek, r. W. (1950). High-yield histologie (2e ed.)). Philadelphia, Pennsylvania: Lippinott Williams & Wilkins.
- Fox, s. Je. (2006). Menselijke fysiologie (9e ed.)). New York, VS: McGraw-Hill Press.
- Gartner, l., & Hiatt, J. (2002). Histologie Atlas Tekst (2e ed.)). Mexico D.F.: McGraw-Hill Inter-Amerikaanse editors.
- Guyton, a., & Hall, J. (2006). Leerboek van medische fysiologie (11e ed.)). Elsevier Inc.
- Johnson, K. (1991). Histologie en celbiologie (2e ed.)). Baltimore, Maryland: The National Medical Series for Independent Study.
- Ross, m., & Pawlina, W. (2006). Histologie. Een tekst en atlas met gecorreleerde cel- en moleculaire biologie (5e ed.)). Lippinott Williams & Wilkins.
- « Compacte botkenmerken, structuur, functies
- IJzeroxide (II) structuur, nomenclatuur, eigenschappen, gebruik »