

Reninstructuur, productie, secretie, functies
- 797
- 91
- Aaron Okuneva
De Renina, Ook bekend als angiotensinogease, is het een aspartylprotease dat belangrijke implicaties heeft bij elektrolythomeostase en bloeddrukcontrole bij zoogdieren.
Dit eiwit wordt uitgescheiden van de nier naar de bloedsomloopstroom en is verantwoordelijk voor het verhogen van de bloeddruk bij experimentele dieren wanneer nierextracten worden geïnjecteerd.
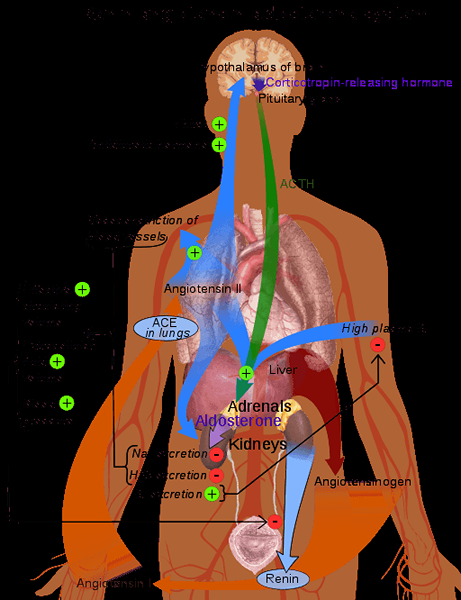
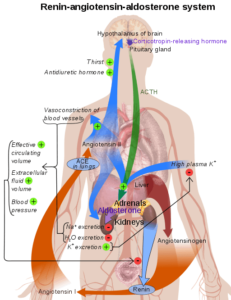 Representatief schema van het renine-angiotensinesysteem in het menselijk lichaam (Bron: Mikael Häggström [Public Domain] via Wikimedia Commons)
Representatief schema van het renine-angiotensinesysteem in het menselijk lichaam (Bron: Mikael Häggström [Public Domain] via Wikimedia Commons) Omdat het een stof is die door een stof wordt geproduceerd en op de circulatie wordt uitgescheiden met een doelwit, weg van de productielocatie, wordt Renin beschouwd als een hormoon.
Hormonen kunnen eiwitten of polypeptiden zijn, steroïde oorsprong hebben of afkomstig zijn van aminozuur tyrosine. Renine is een hormoon van eiwitkarakter en de katalytische werking ervan impliceert de enzymatische clivaje van andere eiwitten (het is een protease).
Dit hormoon werd ontdekt aan het einde van de jaren 1890. Het was echter niets anders dan eind 1990 dat zijn fysiologische oorsprong en moleculaire structuur exact werden bepaald.
[TOC]
Structuur
Menselijke renine is een glycoproteïne met enzymatische activiteit en een molecuulgewicht van iets meer dan 37 kDa. Het molecuul wordt gevormd door twee domeinen gescheiden door een diepe spleet waarbinnen de actieve site zich bevindt.
Beide domeinen van renine zijn vergelijkbaar in volgorde en zijn voornamelijk samengesteld uit β-vergulde vellen.
Verschillende analyse van de sequentie van dit eiwit onthullen dat het meer dan 30 basis aminozuurafval heeft, waaronder verschillende arginines, lysines en histidines.
Bovendien is het bekend dat er gedurende de structuur hydrofoob centrum en grote hydrofiele oppervlakken zijn die stabiliteitseiwit in verschillende contexten bieden.
De actieve plaats van het enzym bevindt zich in de gespleten gevormde door de twee domeinen en de essentiële aminozuren voor katalyse zijn twee asparaginezuurafval in posities 38 en 226, daarom is dit een "aspartil" -protease.
Productie
De renine komt voor in het juxtaglomerulaire systeem van de nier, een gespecialiseerde structuur die wordt aangetroffen in de contactplaats tussen de distale gevormde tubulus en zijn oorsprong glomerulus.
Dit apparaat bestaat uit drie componenten: de korrelige cellen, de extraglomerulaire mesangiale cellen en de dichte macula.
Dichte macula
De dichte macula wordt gevormd door een rij kubieke epitheelcellen die zeer compact zijn gekoppeld die de buis in de contactlocatie met de glomerulus bekleedde en dat wordt beschouwd als het begin van de distale voorgevormde tubulus.
Kan u dienen: Ribulosa: kenmerken, structuur en functiesMesangiale cellen
Extraglomerulaire mesangiale cellen vormen een driehoekig gebied tussen de afferente arteriola, de efferente arteriola en de dichte macula, ze worden beschouwd als een uitbreiding van de glomerulaire mesangiale cellen. Ze worden ook agranulaire cellen genoemd.
Korrelige cellen
Granulaire cellen worden juxtaglomerulaire cellen genoemd en bevinden zich op de wanden van afferente en efferente arteriolen en op het gebied van extraglomerulaire mesangiale cellen.
Deze korrelige cellen ontvangen deze denominatie voor de aanwezigheid van secretie -korrels in hun cytoplasma. Korrels die renine bevatten, evenals een voorloper van renine, pro-reein, die wordt gevormd uit pre-pr-renine.
Pre-Pro-Renina is een prehormoon dat 406 aminozuren bij mensen heeft. Dit prehormoon ondergaat een post-translationele proteolytische clivaje, die een reeks van 23 afval verliest aan zijn terminale amino-uiteinde.
De Cliving van Pre-Pro-Renine zet het om in pro-Reign, 383 aminozuren. De achterste clivaje van een andere sequentie aan het N-terminale uiteinde van pro-afwisseling is wat de vorming van de renine stuurt, een actief protease van 340 aminozuren.
Zowel pro-rrenine als renine kunnen worden uitgescheiden op circulatie, maar zeer weinig pro-afwisseling wordt omgezet in actieve renine in dit bindweefsel. De enzymen die de leiding geven aan de conversie van renine -renine staan bekend als Calicreins en Catopsins.
Zodra de renine is uitgescheiden op de circulatie, heeft deze een halfwaardetijd van maximaal 80 minuten en is de secretie sterk gereguleerd.
Naast de nier kan renine worden geproduceerd door andere weefsels of organen zoals testikels, eierstokken, de wanden van de arteriolen, de bijnierschors, de hypofyse, de hersenen, de hersenen, de vruchtwatervloeistof en anderen.
Hoewel het voor veel dieren van toepassing is, tonen studies met de verwijdering van de nieren aan dat de renine -activiteit in omloop drastisch daalt tot niveaus die zeer dicht bij nul zijn.
Afscheiding
De secretie van de renine neemt toe door een reeks stimuli die verschijnen wanneer het volume van de extracellulaire vloeistof afneemt, wanneer de bloeddruk afneemt of wanneer de sympathische activiteit in nierzenuwen toeneemt.
Verschillende factoren die verband houden met de regulering van reninesecretie zijn beschreven:
- De nierperfusiedruk gedetecteerd door de baroreceptoren (stretchreceptoren) van de afferente arteriola
- Veranderingen in het volume en de samenstelling van de vloeistof die de dichte macula bereikt
- Nier sympathische zenuwen activiteit
- Prostaglandins
- Het natriuretische atriale peptide.
Het baroreceptormechanisme van de afferente arteriola veroorzaakt een afname van de reninesecretie wanneer een toename van de druk van de afferente arteriola op het niveau van het juxtaglomerulaire apparaat optreedt. De secretie neemt toe wanneer de baroreceptoractiviteit afneemt wanneer de druk daalt.
Kan u van dienst zijn: Stroma (histologie)Een andere sensor gerelateerd aan de regulering van reninesecretie is te vinden in de dichte macula. Hoe hoger de reabsorptiesnelheid van Na+ en Cl- en de concentratie van deze elektrolyten in de vloeistof die de dichte macula bereikt, hoe lager de secretie van renine en vice versa.
De toename van de activiteit van de renale sympathische zenuwactiviteit, evenals circulerende catecholamines door norepinefrine die vrijkomt bij sympathische terminaties in juxtaglomerulaire cellen, verhoogt de secretie van de renine.
Prostaglandines, met name Prostacicline.
Angiotensine II, vanwege een negatief feedback -effect, remt de secretie van de renines door direct effect op korrelige cellen. Een ander hormoon zoals vasopressine remt reninesecretie.
Het natriuretische atriale peptide (PNA), dat optreedt in de cardiale atriale spier, remt de secretie van de renines.
Het extra effect van alle stimulerende en remmende factoren is wat de reninesecretie bepaalt. Renine wordt uitgescheiden in nierbloed en laat vervolgens de nieren achter om door het lichaam te circuleren. Er blijft echter een kleine hoeveelheid renine over in niervloeistoffen.
Functie
Renin is een enzym dat op zichzelf geen vasoactieve functies heeft. De enige bekende functie van renine is om angiotensinogeen aan het amino -terminale uiteinde te snijden, een onthoofde genaamd angiotensine i te genereren.
Angiotensinogeen is een glycoproteïne van de groep α2 -globulines gesynthetiseerd door de lever en is aanwezig in het circulerende bloed.
Aangezien angiotensine I een zeer slechte vasopressor-activiteit heeft en het moet door een ander protease "stroomafwaarts" worden verwerkt, neemt de renine deel aan de initiële stappen van bloeddrukregulatie, in een systeem dat bekend staat als renine-angiotensine.
Angiotensin II heeft een zeer korte halfwaardetijd (tussen 1 en 2 minuten). Het wordt snel gemetaboliseerd door verschillende peptidasen die het fragmenteren en sommige van die fragmenten, zoals angiotensine III, behouden wat vasopressor -activiteit.
De algemene functies van het renine -angiotensinesysteem zijn meervoudig en kunnen als volgt worden samengevat:
- Arteriolaire vernauwing en toename van systolische en diastolische druk. Angiotensine II is vier tot acht keer krachtiger dan norepinefrine voor deze functie.
Kan u dienen: Hyracotherium: kenmerken, voeding, soort, reproductie- Verhoging van aldosteronsecretie als gevolg van het directe effect van angiotensine II op de bijnierschors. Het renine-angiotensinesysteem is de belangrijkste regulator van aldosteron-secretie.
- Het vergemakkelijkt norepinefrine-secretie vanwege direct effect op sympathische postgangliones.
- Het beïnvloedt mesangiale celcontractie, wat de glomerulaire filtratiesnelheid vermindert en, vanwege het directe effect op nierbuisjes, de natriumreabsorptie verhoogt.
- Op hersenniveau vermindert dit systeem de gevoeligheid van de baroreceptorreflex die het angiotensine II vasopressor -effect verbetert.
- Angiotensine II stimuleert de waterinname door dorstmechanismen te bevorderen. Verhoogt de secretie van vasopressine en ACTH -hormoon.
Gerelateerde pathologieën
Het renine-angiotensinesysteem speelt daarom een belangrijke rol in hypertensieve pathologieën, met name die van nieroorsprong.
Dit is hoe de vernauwing van een van de nierslagaders aanhoudende hypertensie genereert die kan worden teruggedraaid als de ischemische (defecte) nier wordt geëxtraheerd of de nierarteriële vernauwing wordt vrijgegeven in de tijd.
Een toename van de renineproductie wordt in het algemeen geassocieerd met de unilaterale vernauwing van de nierslagader die een van de nieren verbindt, wat hypertensie veroorzaakt. Deze klinische toestand kan te wijten zijn aan aangeboren defecten of andere afwijkingen van de niercirculatie.
De farmacologische manipulatie van dit systeem, naast het gebruik van blokkers van angiotensine II -receptoren, zijn de fundamentele hulpmiddelen voor de behandeling van arteriële hypertensie.
Arteriële hypertensie is een stille en progressieve ziekte die een groot deel van de wereldbevolking treft, vooral 50 jaar oude volwassenen.
Referenties
- Akahane, k., Umeyama, h., Nakagawa, s., Moriguchi, ik., Hirose, s., Iizuka, k., & Murakami, J. (1985). Driedimensionale structuur van menselijke renine. Hypertensie, 7(1), 3-12.
- Davis, J., & Freeman, r. (1976). Mechanismen die Renin DeLease reguleren. Fysiologische beoordelingen, 56(1), 1-56.
- Guyton, a., & Hall, J. (2006). Leerboek van medische fysiologie (11e ed.)). Elsevier Inc.
- Hackenthal, e., Paul, m., Ganten, D., & Taugner, r. (1990). Morfologie, fysiologie en moleculaire biologie van Renin Secret. Fysiologische beoordelingen, 70(4), 1067-1116.
- Morris, B. (1992). Biologie van Renin Molecular. I: Gene- en eiwitstructuur, synthese en verwerking. Journal of Hypertension, 10, 209-214.
- Murray, r., Bender, D., Botham, k., Kennelly, p., Rodwell, V., & Weil, p. (2009). Harper's geïllustreerde biochemie (28e ed.)). McGraw-Hill Medical.
- West, J. (1998). Fysiologische basis van de medische praktijk (12ava ed.)). Mexico D.F.: Pan -Aricaans medisch redactioneel.