

Fylogenetische boomtypen en -kenmerken, voorbeelden
- 4005
- 459
- James Dach
A fylogenetische boom Het is een wiskundige grafische weergave van geschiedenis en voorouderrelaties - afstammelingen van groepen, populaties, soorten of een andere taxonomische categorie. Theoretisch kunnen alle fylogenetische bomen worden gegroepeerd in de levensboom, die de universele boom vormen.
Deze grafische representaties hebben een revolutie teweeggebracht in de studie van de evolutionaire biologie, omdat ze het mogelijk maken om een soort te vestigen en te definiëren, verschillende evolutionaire hypothesen (zoals endosimbiotische theorie) te testen, waarbij de oorsprong van ziekten (zoals HIV), enz.
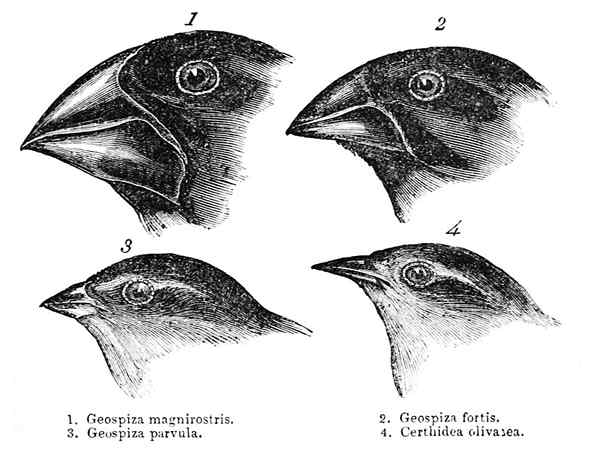
Bron: John Gould (14.Sep.1804 - 3.Februari.1881) [Public Domain] Bomen kunnen worden herbouwd met behulp van morfologische of moleculaire karakters, of beide. Op dezelfde manier zijn er verschillende methoden om ze te bouwen, de meest voorkomende is de cladistische methodologie. Dit wil gedeelde afgeleide tekens identificeren, bekend als Synapomorphia.
[TOC]
Kenmerken
Een van de principes ontwikkeld door Charles Darwin vormt de gemeenschappelijke afkomst van alle levende organismen - dat wil zeggen dat we allemaal een afgelegen voorouder delen.
In "Het ontstaan van soorten" Darwin verhoogt de metafoor van de "levensboom". Gebruik in feite een hypothetische grafische boom om uw idee te ontwikkelen (merkwaardig genoeg is dit de enige illustratie van de Oorsprong)).
De weergave van deze metafoor is wat we kennen als fylogenetische bomen, waardoor we de geschiedenis en relaties van een concrete groep organismen grafisch kunnen vertonen.
Anatomie van een fylogenetische boom
In fylogenetische bomen kunnen we de volgende delen onderscheiden - doorgaan met botanische analogie:
Takken: De boomlijnen worden "takken" genoemd, en deze vertegenwoordigen de studiepopulaties in de tijd. Afhankelijk van het type boom (zie later), kan de lengte van de tak al dan niet een betekenis hebben.
Op het puntje van de takken vinden we de organismen die we willen evalueren. Dit kunnen entiteiten zijn die momenteel levend zijn, of uitgestorven wezens. De soort zou de bladeren van onze boom zijn.
Wortel: De wortel is de meest voorouderlijke tak van de boom. Sommigen hebben het en worden gewortelde bomen genoemd, terwijl anderen het niet hebben.
Knooppunten: De vertakkingspunten van de takken in twee of meer lijnen worden knooppunten genoemd. Het punt vertegenwoordigt de meest recente gemeenschappelijke voorouder van de afstammelingen (merk op dat deze voorouders hypothetisch zijn).
Het bestaan van een knooppunt impliceert een speciatie -gebeurtenis - creatie van nieuwe soorten. Hierna volgt elke soort zijn evolutionaire koers.
Aanvullende terminologie
Naast deze drie basisconcepten zijn er andere noodzakelijke termen met betrekking tot fylogenetische bomen:
Politiek: Wanneer het in een fylogenetische boom meer dan twee takken in een knooppunt presenteert, wordt gezegd dat er een politieke is. In deze gevallen is de fylogenetische boom niet volledig opgelost, omdat de relaties tussen de betrokken organismen niet duidelijk zijn. Dit gebeurt meestal vanwege het gebrek aan gegevens en kan alleen worden opgelost wanneer een onderzoeker meer ophoopt.
Kan u dienen: Tetrosas: kenmerken, erythrosa, synthese, derivatenExterne groep: In fylogenetische thema's is het gebruikelijk om te luisteren naar het concept van externe groep - ook wel genoemd Outgroep. Deze groep is geselecteerd om de boom te kunnen rooten. Het moet worden gekozen als een taxon dat eerder de studiegroep heeft uiteengezet. Als ik bijvoorbeeld echinodermos studeer, kunt u gebruiken Outgroep naar de ascidia.
Jongens
Er zijn drie basistypen bomen: cladogrammen, additieve bomen en ultra -gebaseerde bomen.
Clayogrammen zijn de eenvoudigste bomen en vertonen de relatie van organismen in termen van gemeenschappelijke afkomst. Informatie van dit type boom ligt in vertakkingspatronen, omdat de grootte van de takken geen extra betekenis heeft.
Het tweede type boom is het additief, ook wel metrische bomen of philogrammen genoemd. De lengte van de takken is gerelateerd aan de hoeveelheid evolutionaire verandering.
Ten slotte hebben we ultrametrische bomen of dendogrammen, waarbij alle tips van de bomen zich op dezelfde afstand bevinden (wat niet gebeurt in het Philogram, waar een tip hieronder of hoger kan verschijnen dan uw partner). De lengte van de tak is gerelateerd aan de evolutietijd.
De keuze van de boom is direct gerelateerd aan de evolutionaire vraag die we willen beantwoorden. Als we bijvoorbeeld alleen om relaties tussen individuen geven, zal een cladogram voldoende zijn voor de studie.
Meest voorkomende fouten bij het lezen van fylogenetische bomen
Hoewel fylogenetische bomen meestal afbeeldingen zijn van breed gebruik in de evolutiebiologie (en in de algemene biologie), zijn er veel studenten en professionals die de boodschap verkeerd begrijpen dat deze graphics - blijkbaar eenvoudig - van plan zijn om aan de lezer te leveren.
Er is geen romp
De eerste fout is om ze lateraal te lezen, ervan uitgaande dat evolutie vooruitgang impliceert. Als we het evolutieproces correct begrijpen, is er geen reden om te denken dat links de voorouderlijke en juiste soorten de meest gevorderde soorten zijn.
Hoewel boom botanische analogie erg nuttig is, is er een punt waar het niet langer zo nauwkeurig is. Er is een cruciale structuur van de boom die niet aanwezig is in de boom: de romp. In fylogenetische bomen vinden we geen enkele hoofdtak.
In het bijzonder konden sommige mensen de mens beschouwen als het "ultieme doel" van evolutie, en daarom de soort Homo sapiens Het moet altijd worden gelokaliseerd als een laatste entiteit.
Kan u dienen: tumornecrosefactor (TNF): structuur, actiemechanisme, functieDeze visie is echter niet congruent met evolutionaire principes. Als we begrijpen dat fylogenetische bomen mobiele elementen zijn, kunnen we de Homo In elke terminale positie van de boom, omdat dit kenmerk niet relevant is in de weergave.
De knooppunten kunnen roteren
Een essentieel kenmerk dat we van fylogenetische bomen moeten begrijpen, is dat ze niet -statische afbeeldingen vertegenwoordigen.
In hen kunnen al deze takken roteren - op dezelfde manier als een mobiel het kan. We bedoelen niet dat de takken naar onze inboorlingen kunnen worden verplaatst, omdat sommige bewegingen de verandering van het patroon zouden impliceren of Topologie Van de boom. Wat we kunnen roteren zijn de knooppunten.
Om de boodschap van een boom te interpreteren, moeten we ons niet concentreren op het puntje van de takken, we moeten het doen op de vertakkingspunten, die het belangrijkste aspect van de afbeelding zijn.
Bovendien moeten we rekening houden met dat er verschillende manieren zijn om een boom te tekenen. Vaak hangt het af van de stijl van het boek of het tijdschrift en de veranderingen in de vorm en positie van de takken hebben geen invloed op de informatie die ze willen overbrengen.
We kunnen het bestaan van voorouderlijke of "oude" soorten niet afleiden
Als we naar soorten gaan verwijzen huidig We moeten geen voorouderlijke connotaties toepassen. Als we bijvoorbeeld denken aan de relaties tussen chimpansees en mensen, kunnen we ten onrechte begrijpen dat chimpansees voorouderlijk zijn met betrekking tot onze afkomst.
De gemeenschappelijke voorouder van chimpansees en mensen was echter geen van beide. Om te denken dat chimpansee voorouderlijk is, zou het zijn om aan te nemen dat de evolutie ervan stopte zodra beide lijnen waren gescheiden.
In navolging van dezelfde logica van deze ideeën, geeft een fylogenetische boom niet aan of er jonge soorten zijn. Aangezien allelische frequenties constant zijn en er in de loop van de tijd nieuwe personages veranderen, is het moeilijk om de leeftijd van een soort te bepalen en, zeker, een boom geeft ons dergelijke gegevens niet.
De "verandering van allelische frequenties in de loop van de tijd" is de manier waarop populaties genetica evolutie definiëren.
Ze zijn onveranderlijk
Bij het observeren van een fylogenetische boom moeten we begrijpen dat deze grafiek gewoon een hypothese is die is gegenereerd uit concreet bewijs. Als we meer tekens aan de boom toevoegen, wijzigt het de topologie.
De expertise van wetenschappers bij het kiezen van de beste personages die het toestaan de relaties van organismen in kwestie op te helderen, is de sleutel. Bovendien zijn er zeer krachtige statistische hulpmiddelen waarmee onderzoekers bomen kunnen evalueren en de meest plausibele kunnen kiezen.
Kan u van dienst zijn: proline: kenmerken, structuur, functies, voedselVoorbeelden
The Three Domains of Life: Archaea, Bacteria en Eukarya
In 1977 stelde onderzoeker Carl Woese voor om levende organismen te groeperen in drie domeinen: Archaea, Bacteria en Eukarya. Dit nieuwe classificatiesysteem (voorheen waren er slechts twee categorieën, Eukaryota en Prokaryota) was gebaseerd op de ribosomale ribmoleculaire marker.
Bacteriën en eukaryoten zijn algemeen bekende organismen. Archaeas worden meestal verward met bacteriën. Ze verschillen echter diep in de structuur van hun celcomponenten.
Hoewel het microscopische organismen zoals bacteriën zijn, zijn de leden van het Archaea -domein daarom meer gerelateerd aan eukaryoten - omdat ze een voorouder in een dichterbijzijnde delen delen.
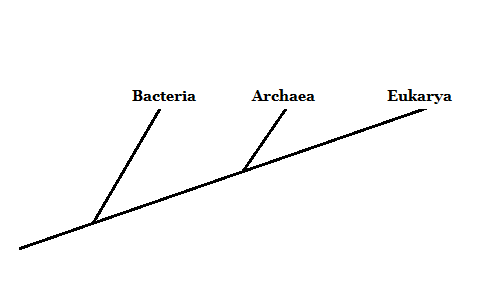 Bron: Opgesteld door Mariana Gelambi.
Bron: Opgesteld door Mariana Gelambi. Filogenese van primaten
Binnen de evolutiebiologie is een van de meest controversiële kwesties de evolutie van de mens. Voor de tegenstanders van deze theorie is er geen logica voor een evolutie op basis van een Simiesco -voorouder die aanleiding gaf tot de huidige man.
Een belangrijk concept is om te begrijpen dat we niet evolueren van de huidige apen, maar dat we een gemeenschappelijke voorouder met hen delen. In de boom van apen en mensen benadrukt het dat wat we kennen als "aap" geen geldige monofiletische groep is, omdat het de mens uitsluit.
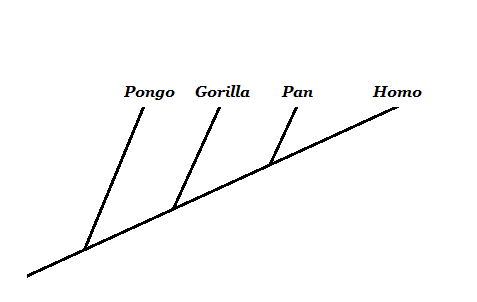 Bron: Opgesteld door Mariana Gelambi.
Bron: Opgesteld door Mariana Gelambi. Fylogenese van cetartiodactilos (cetartiodactyla)
Evolutionair gezien vertegenwoordigden de walvisachtigen een groep gewervelde dieren waarvan de relaties met de rest van hun metgezellen zoogdieren niet erg duidelijk waren. Morfologisch gezien hebben walvissen, dolfijnen en andere leden weinig overeenkomsten met de rest van de zoogdieren.
Momenteel, dankzij de studie van verschillende morfologische en moleculaire karakter.
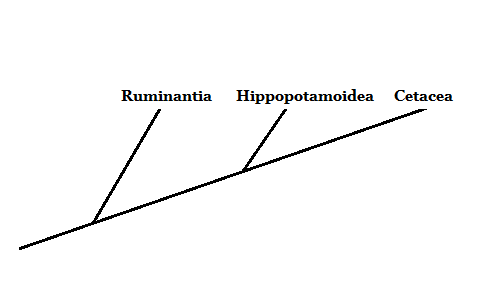 Bron: Opgesteld door Mariana Gelambi.
Bron: Opgesteld door Mariana Gelambi. Referenties
- Baum, D. NAAR., Smith, s. D., & Donovan, s. S. (2005). De boom-denkende uitdaging. Wetenschap, 310(5750), 979-980.
- Curtis, h., & Barnes, n. S. (1994). Uitnodiging voor de biologie. Macmillan.
- Hall, B. K. (ED.)). (2012). Homologie: de hiërarchische basis van vergelijkende biologie. Academische pers.
- Hickman, c. P., Roberts, l. S., Larson, a., Ober, W. C., & Garrison, c. (2001). Geïntegreerde priorms van zoölogie. McGraw-Hill.
- Kardong, K. V. (2006). Gewervelde dieren: vergelijkende anatomie, functie, evolutie. McGraw-Hill.
- Kliman, r. M. (2016). Encyclopedie van evolutiebiologie. Academische pers.
- Losos, J. B. (2013). The Princeton Guide to Evolution. Princeton University Press.
- Pagina, r. D., & Holmes, en. C. (2009). Moleculaire evolutie: een fylogenetische benadering. John Wiley & Sons.
- Rijst, s. NAAR. (2009). Encyclopedie van evolutie. Informeren publiceren.
- Starr, c., Evers, c., & Starr, L. (2010). Biologie: concepten en toepassingen zonder fysiologie. Cengage leren.
- « Euler -methode voor wat het gebruik is van procedure en oefeningen
- De 10 moeilijkste engineering om te studeren »