

Nucleaire poriënkenmerken, functies, componenten
- 3550
- 585
- Cecil Graham
Hij nucleaire porie (Van het Grieks, poriën = stap of doorvoer) is de "deur" van de kern die het grootste deel van het transport tussen nucleoplasma en cytoplasma mogelijk maakt. De nucleaire porie sluit zich aan bij de interne en externe membranen van de kern om kanalen te vormen, die dienen voor het transport van eiwitten en RNA.
Het woord poro weerspiegelt niet de complexiteit van de structuur in kwestie. Daarom heeft het de voorkeur om te verwijzen naar het nucleaire poriecomplex (CPN), in plaats van nucleaire porie. De CPN kan veranderingen in zijn structuur ervaren tijdens transport of toestanden van de celcyclus.
Onlangs is ontdekt dat nucleoporines, eiwitten die de CPN vormen, een belangrijke rol spelen bij het reguleren van genexpressie. Dus wanneer mutaties optreden die de functie van nucleoporines beïnvloeden, worden pathologieën geproduceerd bij mensen, zoals auto -immuun, cardiomyopathieën, virale infecties en kanker.
[TOC]
Kenmerken
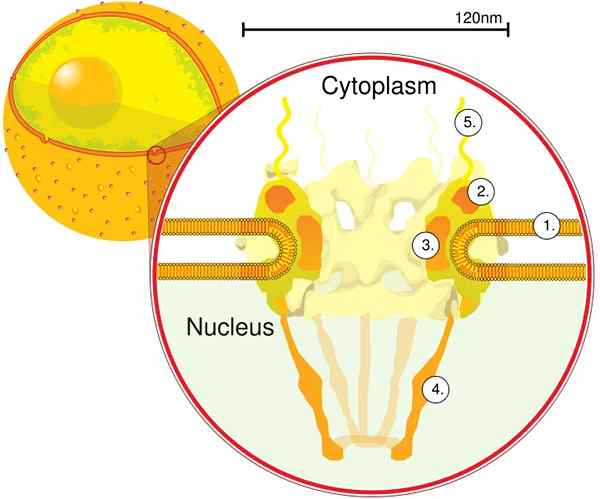
Door elektronentomografie werd vastgesteld dat de CPN een dikte van 50 nm heeft, een externe diameter tussen 80 en 120 nm en een interne diameter van 40 nm. Grote moleculen zoals de grote subeenheid van ribosomen (PM 1590 kDa) kunnen via de CPN buiten de kern worden geëxporteerd. Naar schatting zijn er tussen 2000 en 4000 CPN per kern.
Het molecuulgewicht van een individuele CPN is ongeveer tussen 120 en 125 MDa (1 MDA = 106 Da) in gewervelde dieren. De CPN is daarentegen kleiner in gisten, waarin hij ongeveer 60 MDA heeft. Ondanks de enorme grootte van de CPN zijn nucleoporines sterk bewaard in alle eukaryoten.
Translocalisatie door de CPN is een snel proces, waarvan de snelheid 1000 translocaties is/tweede. De CPN bepaalt echter niet de richting van de transportstroom zelf.
Dit hangt af van de RangTP -gradiënt, die groter is in de kern dan in de cytosol. Deze gradiënt wordt gehandhaafd door een guanine -uitwisselaarfactor die liep.
Tijdens de celcyclus ervaren de CPNS -cycli assemblage en verdeeldheid. De montage vindt plaats op de interface en onmiddellijk na mitose.
Functie
Ribonucleïnezuur (klein nucleair RNA, messenger -RNA, overdracht RNA), eiwitten en ribonucleoproteïnen (RNP) moeten actief worden getransporteerd via CPN. Dit betekent dat de energie van ATP- en GTP -hydrolyse vereist is. Elk molecuul wordt op een specifieke manier getransporteerd.
Over het algemeen zitten RNA -moleculen boordevol eiwitten die RNP -complexen vormen, die op deze manier worden geëxporteerd. Eiwitten die actief naar de kern worden getransporteerd, moeten daarentegen een sequentieteken van locatie hebben in de kern (SLN), bezitter van aminozuurafval met positieve belasting (bijvoorbeeld KKKRK).
Kan u van dienst zijn: mesosoomEiwitten die naar de kern worden geëxporteerd, moeten een exportsignaal hebben voor de kern (NES) die rijk is aan leucine -aminozuur.
Naast het faciliteren van transport tussen de kern en cytoplasma, zijn CPN's betrokken bij de organisatie van de chromatine, genexpressieregulatie en DNA -herstel. Nucleoporines (NUP's) bevorderen transcriptieactivering of repressie, ongeacht de celproliferatietoestand.
In gisten worden de nupen gevonden in de CNP van de nucleaire wikkeling. In de metazoaries zijn ze binnen. Ze voeren dezelfde functies uit in alle eukaryoten.
Import van stoffen
Via de CPN is er passieve verspreiding van kleine moleculen in beide richtingen en actief transport, eiwitimport, export van RNA en ribonucleoproteïnen (RNP's) en de bidirectionele moleculen shuttle. De laatste omvat RNA, RNP en eiwitten die betrokken zijn bij de signalering, biogenese en vervanging.
De invoer van eiwitten in de kern vindt plaats in twee stappen: 1) eiwitbinding aan de cytoplasmatische zijde van de CPN; 2) ATP -afhankelijke translocalisatie door de CPN. Dit proces heeft ATP -hydrolyse en de uitwisseling van GTP/BBP tussen de kern en het cytoplasma nodig.
Volgens een transportmodel beweegt het ontvangende eiwitcomplex door de unie, dissociatie en unie opnieuw naar de herhaalde FG-sequenties van de nucleoporines. Op deze manier beweegt het complex van de ene nucleoporine naar de andere binnen de CPN.
Export van stoffen
Is vergelijkbaar met import. Ran GTPase legt directionaliteit op aan transport door CNP. Ran is een moleculaire schakelaar met twee conformationele toestanden, afhankelijk van of het is gekoppeld aan de BBP of GTP.
Twee specifieke regulerende eiwitten veroorzaken de conversie tussen de twee toestanden: 1) cytosolische GTPase-activerende eiwit (GAP), die GTP-hydrolyse produceert en aldus RAN-GTP converteert naar RAN-GDP; en 2) nucleaire guanina-uitwisselingsfactor (GEF), die de uitwisseling van het bbp bevordert door GTP en converteert Ran-GDP naar Ran-GTP.
Cytosol bevat voornamelijk RAN-GDP. De kern bevat voornamelijk RAN-GTP. Deze gradiënt van de twee conformationele vormen van RAN richt het transport in het juiste adres.
De invoer van de ontvanger, samen met de positie, wordt door de Unie vergemakkelijkt naar de herhalingen-FG. Als u de nucleaire kant van de CNP bereikt, sluit Ran-GTP zich aan bij de ontvanger om uw positie vrij te geven. RAN-GTP creëert dus de richting van het importproces.
Nucleaire export is vergelijkbaar. Ran-GTP in de kern bevordert echter de unie van de positie tot de exportontvanger. Wanneer de exportontvanger door de porie naar de cytosol beweegt, voldoet deze aan de ran-gap, die GTP-hydrolyse aan bbp induceert. Ten slotte wordt de ontvanger vrijgelaten uit zijn positie en ran-gdp in de cytosol.
Kan u van dienst zijn: polymorfonucleaire leukocytenRNA -transport
De export van sommige soorten RNA is vergelijkbaar met de export van eiwitten. ARNT en RNAN's (kleine nucleair) gebruiken bijvoorbeeld de RangTP -gradiënt en worden door middel van de CPN getransporteerd. De export van volwassen ribosomen is ook afhankelijk van de rangtp -gradiënt.
Het mRNA wordt op een heel andere manier geëxporteerd dan de eiwitten en andere RNA's. Voor zijn export vormt het mRNA een RNP -messenger (RNPM) -complex, waarin een RNM -molecuul is omgeven door honderden eiwitmoleculen. Deze eiwitten hebben de verwerking, Aftappen, Empalme en Polyadentilly van MNA.
De cel moet in staat zijn om onderscheid te maken tussen RNPM met volwassen RNM en RNPM met onrijpe mRNA. Het mRNA, dat het RPNM -complex vormt, zou topologieën kunnen overnemen die moeten worden gerenoveerd voor transport. Voordat RNPM de CPN binnengaat, treedt een controlestap uit door tramp- en exosoomeiwitcomplexen op.
Wanneer volwassen RNPM wordt geassembleerd, wordt RPNM door het kanaal getransporteerd door middel van een transportontvanger (NXF1-NXT1). Deze ontvanger heeft ATP -hydrolyse nodig (geen RangTP -gradiënt) om de directionaliteit van de RNPM vast te stellen, die het cytoplasma zal bereiken.
Het nucleaire poriecomplex en de controle van genexpressie
Sommige studies geven aan dat CPN -componenten de regulatie van genetische expressie kunnen beïnvloeden door de controle van de chromatinestructuur en de toegankelijkheid ervan tot transcriptiefactoren.
In eukaryoten van recentere evolutie bevindt heterochromatine zich bij voorkeur aan de periferie van de kern. Dit gebied wordt onderbroken door euchromatinekanalen, die worden gehandhaafd door de CPN -nucleaire mand. De nucleaire mandassociatie met euchromatine is gerelateerd aan gentranscriptie.
De activering van transcriptie op het CPN -niveau impliceert bijvoorbeeld de interactie van de componenten van de nucleaire mand met eiwitten zoals de histonacetyltransferase -saga en RNA -exportfactoren.
De nucleaire mand is dus een platform voor tal van onderhoudsgenen (Huishouden) sterk getranscribeerde en genen sterk geïnduceerd door veranderingen in omgevingscondities.
Het nucleaire porie- en virologiecomplex
Virale eukaryotcelinfectie hangt af van CPN. In elk geval van virale infectie hangt het succes ervan af van DNA, RNA of RPN dat door de CPN gaat om zijn uiteindelijke doel te bereiken, wat de replicatie van het virus is.
Kan u van dienst zijn: cytoplasmatische insluitsels: wat zijn, kenmerken, functiesAPE -virus 40 (SV40) is een van de meest bestudeerde modellen geweest om de functie van de CPN in translocalisatie in de kern te onderzoeken. Dit komt omdat SV40 een klein genoom heeft (5.000 bases).
Er is aangetoond dat het transport van het virus -DNA -eiwiteiwitten van het virus is vergemakkelijkt, die het virus beschermen totdat de kern bereikt.
Componenten
De CPN is ingebed in de nucleaire envelop en bestaat uit tussen ongeveer 500 en 1000 NUPTS. Deze eiwitten zijn georganiseerd in structurele subcomplexen of modules, die met elkaar interageren.
De eerste module is een centrale component, of ring, in de porie in de vorm van een zandklok, die wordt beperkt door een andere 120 nm diameter ring aan beide zijden, intranucleair en cytoplasmatisch. De tweede module zijn de kern- en cytoplasma -ringen (elk met een diameter van 120 nm) die zich rond de zandwaatvormige component bevinden.
De derde module zijn acht filamenten die worden geprojecteerd vanuit de 120 nm ring in de nucleoplasma en vormt een mandvormige structuur. De vierde module bestaat uit de filamenten die naar de zijkant van het cytoplasma worden geprojecteerd.
Het y -vormige complex, bestaande uit zes Nups en SEH 1 en SEC 13 -eiwitten, is het grootste en best gekarakteriseerde complex van de CNP. Dit complex is de essentiële eenheid die deel uitmaakt van de CPN -steiger.
Ondanks de lage gelijkenis tussen de Nups -sequenties, is de CPN -steiger zeer bewaard in alle eukaryoten.
Referenties
- Beck, m., Pijn doen, e. 2016. Het nucleaire poriecomplex: de functie ervan begrijpen door structureel inzicht. Nature Reviews, Molecular Cell Biology, DOI: 10.1038/nrm.2016.147.
- Ibarra, een., Hetzer, m.W. 2015. Nucleaire porie -eiwitten en de controle van genoomfuncties. Genen en ontwikkeling, 29, 337-349.
- Kabachinski, g., Schwartz, T.OF. 2015. De nucleaire poriecomplex - structuur en functie in één oogopslag. Journal of Cell Science, 128, 423-429.
- Knocknhauer, K.EN., Schwartz, T.OF. 2016. Het nucleaire poriëncomplex als flexibele en dynamische poort. Cell, 164, 1162-1171.
- Ptak, C., Aitchison, J.D., Wozniak, r.W. 2014. Het multifunctionele nucleair door complex: een platform voor het beheersen van genexpressie. Huidige mening over celbiologie, DOI: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Republicatie: het nucleaire poriecomplex - een uitgebreid overzicht van structuur en functie. International Journal of Academic Medicine, 3, 51-59.
- Tran, e.J., Wente, s.R. 2006. Dynamisch nucleair door complexen: leven op de rand. Cell, 125, 1041-1053.