

Vloeibaar mozaïek model
- 3783
- 336
- Aaron Okuneva
We leggen uit wat het vloeistofmozaïekmodel, de structuur van het celmembraan en zijn functies is.
Vloeibare mozaïekmodelstructuur Wat is het vloeibare mozaïekmodel?
Hij vloeibaar mozaïek model Het stelt dat cel- of biomembraanmembranen dynamische structuren zijn die vloeiend zijn van hun verschillende moleculaire componenten, die lateraal kunnen bewegen. Dat wil zeggen, deze componenten zijn in beweging en niet -statisch, zoals eerder geloofde.
Dit model is opgevoed door S. Jonathan -zanger en Garth. L. Nicolson in 1972 en wordt vandaag algemeen aanvaard door de wetenschappelijke gemeenschap. Alle cellen zijn opgenomen door een celmembraan met bijzonderheden in zijn grondwet en functie.
Dit membraan definieert de grenzen van de cel, waardoor verschillen tussen cytosol (of cellulair interieur) en de externe omgeving mogelijk zijn. Het reguleert ook de uitwisseling van stoffen tussen de cel en de buitenkant.
In eukaryotische cellen definiëren interne membranen ook compartimenten en organellen met verschillende functies, zoals mitochondriën, chloroplasten, nucleaire envelop, endoplasmatisch reticulum, Golgi's apparaat, onder andere.
Celmembraanstructuur
Algemeenheid
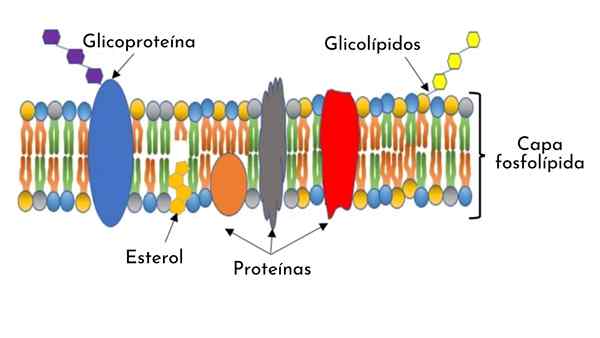
- Het celmembraan bestaat uit een waterdichte structuur tot moleculen en in water oplosbare ionen tussen 7 en 9 nanometer dik. Het wordt waargenomen in elektronische microfotografieën als een dubbele continue en dunne lijn die het celcytoplasma omringt.
- Het is samengesteld uit een fosfolipide dubbellaag, met eiwitten ingebed langs de structuur en op het oppervlak gerangschikt.
- Het bevat koolhydraatmoleculen op beide oppervlakken (intern en extern) en in het geval van eukaryotische cellen van dieren presenteert het ook cholesterolmoleculen afgewisseld in de dubbellager.
Fosfolipide dubbellaag
Fosfolipiden zijn amfipatische moleculen met een hydroopofisch uiteinde -waterfine -en een ander hydrofoob -dat water afstoot-.
De fosfolipide dubbellaag die het celmembraan vormt, heeft hydrofobe (apolaire) ketens die in het membraan zijn gerangschikt en hydrofiele (polaire) extremen zich in de richting van het externe medium.
Het kan u van dienst zijn: Flora en Fauna uit de Verenigde StatenAldus worden de hoofden van fosfaatgroepen fosfolipiden blootgesteld op het buitenoppervlak van het membraan.
Bedenk dat zowel de externe als het interne of cytosol waterig zijn. Dit beïnvloedt de opstelling van de dubbele fosfolipidelaag, met zijn polaire delen interactie met water en zijn hydrofobe delen die de interne membraanmatrix vormen.
Cholesterol
In het membraan van eukaryot -dierlijke cellen zijn er cholesterolmoleculen ingebracht in de hydrofobe staarten van fosfolipiden.
Deze moleculen worden niet gevonden in de membranen van prokaryotische cellen, sommige protisten, planten en schimmels.
Uitgebreide membraan- of transmembraaneiwitten
In de fosfolipide tweewegen.
Deze interageren niet covalent door hun hydrofobe delen, met lipidebilay.
Membraaneiwitconfiguratie
Ze kunnen een eenvoudige configuratie presenteren in de vorm van een Baston, met een alfa -hydrofobe en ingebed alfa -propeller in het membranale interieur, en met uitgebreide hydrofiele delen aan de zijkanten.
Ze kunnen ook een grotere, bolvormige type configuratie presenteren, en met een complexe tertiaire of quaternaire structuur.
De laatste kruist het celmembraan vaak meerdere keren met hun alfa -propellersegmenten herhaald en gerangschikt in zigzag door de lipide dubbellaag.
Poriën in de membranen
Sommige van deze bolvormige eiwitten hebben hydrofiele binnengedeelten, die kanalen of poriën vormen waardoor de uitwisseling van polaire stoffen optreedt van de cel buiten naar de cytosol, en vice versa.
Perifere eiwitten
Op het oppervlak van het cytoplasmatische gezicht van het celmembraan zijn er perifere membraaneiwitten, gekoppeld aan de uitstekende delen van sommige integrale eiwitten.
Het kan u van dienst zijn: APTENIA CORDIFOLIA: Kenmerken, habitat, eigenschappen, teeltDeze eiwitten dringen niet door in de hydrofobe kern van de lipide dubbellaag.
Koolhydraatbedekking
Er zijn koolhydraatmoleculen op beide membraanoppervlakken.
In het bijzonder heeft het buitenoppervlak van het membraan een overvloed aan glucolipiden. Korte koolhydraatketens worden ook waargenomen blootgesteld en covalent bevestigd aan de uitstekende eiwitonderdelen, Glycoproteïnen genoemd.
Vloeibaarheid van celmembraan
Verhouding verzadigde vetzuren versus. onverzadigd
De vloeibaarheid van het membraan hangt voornamelijk af van het aandeel tussen fosfolipiden van verzadigde en niet -verzadigde vetzuren aanwezig.
Deze membranale vloeibaarheid neemt af naarmate het aandeel fosfolipiden van verzadigde vetzuurketens toeneemt, met betrekking tot niet -verzadigde.
Dit komt omdat cohesie groter is tussen lange en eenvoudige ketens van verzadigde vetzuren, met betrekking tot cohesie tussen de korte en onverzadigde ketens van onverzadigde vetzuren.
Een grotere cohesie tussen zijn moleculaire componenten, lagere vloeiendheid zal het membraan presenteren.
Cholesterol
Cholesterolmoleculen interageren via hun stijve ringen met lipide koolwaterstofketens, waardoor de stijfheid van het membraan wordt verhoogd en de permeabiliteit daarvan wordt verminderd.
In de membranen van de meerderheid van de eukaryotische cellen, waar een relatieve hoge cholesterolconcentratie is, voorkomt dit dat koolzuurhoudende ketens lage temperaturen samenvoegen. Naar verwachting zal dit membraan bevriezen bij lage temperaturen.
Bijzonderheden
De verschillende soorten celmembranen vertonen bijzonderheden in hun hoeveelheid en type eiwitten en koolhydraten, evenals in de verscheidenheid van bestaande lipiden.
Deze bijzonderheden worden geassocieerd met specifieke cellulaire functies.
Er zijn niet alleen constitutieve verschillen tussen eukaryot en prokaryotische celmembranen en tussen die van organellen, maar ook tussen regio's van hetzelfde membraan.
Kan u van dienst zijn: SSB -eiwitten: kenmerken, structuur en functiesCelmembraanfuncties
- Het celmembraan scheidt de cel en stelt het in staat een stabiele toestand in de cytosol te behouden, anders dan die van de externe omgeving.
- Door de actieve en passieve regulatie van de doorgang van stoffen (water, ionen en metabolieten) door zichzelf, handhaaft het het noodzakelijke elektrochemische potentieel voor cellulaire werking.
- Hiermee kan de cel reageren op externe middelgrote signalen via chemische membraanreceptoren en biedt verankeringssites voor cytoskeletfilamenten.
- In het geval van eukaryotische cellen neemt het ook deel aan de oprichting van interne compartimenten en organellen met specifieke metabole functies.
Eiwitfunctie in het membraan
Er zijn verschillende membraaneiwitten met specifieke functies, waaronder we kunnen vermelden:
- Enzymen die chemische reacties katalyseren (versnellen).
- Membraanreceptoren die deelnemen aan herkenning en unie voor signaalmoleculen (zoals hormonen).
- Substantie transport eiwitten door het membraan (naar de cytosol en van dit naar de buitenkant van de cel). Deze behouden een elektrochemische gradiënt dankzij iontransport.
Externe koolhydraatafdekkingsfunctie
Koolhydraten of glucolipiden nemen deel aan de hechting van cellen met elkaar en aan het herkenningsproces en interactie van celmembraan met moleculen zoals antilichamen, hormonen en virus.
Referenties
- Bagsover, s. R., Hyams, J. S., Shephard, E. NAAR., Wit, h. NAAR. en Wiedemann, c. G. (2003). Celbiologie, tot korte cursus. Tweede druk. Wiley-Liss.
- Engelman, D. (2005). Zijn meer mozaïek zijn vloeibare membranen. Natuur 438 (7068).
- Raven, J. (2002). Biologie. Zesde editie. MGH.