

Hemoolisin -kenmerken, typen, werkingsmechanismen
- 1158
- 65
- Lonnie Rohan
De Hemoolisine Het is een klein eiwit dat poriën in het celmembraan van erytrocyten en sommige andere cellen van het bloed van zoogdieren veroorzaakt. Het wordt over het algemeen gesynthetiseerd en uitgescheiden door pathogene bacteriën.
Dit eiwit is een van de meest voorkomende microbiële toxines en degene die het beste is bestudeerd. Soms kan het hemolytische anemie veroorzaken, omdat de hoeveelheid kanalen waardoor het celinterieur uitkomt, zelfs cellysis kan veroorzaken.
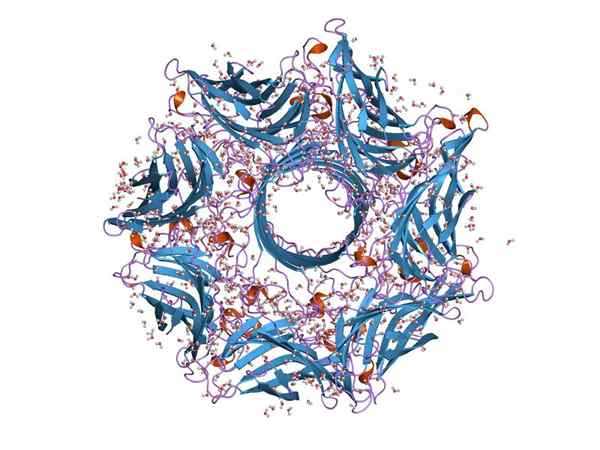
 Moleculaire structuur van een Hemoolisine (Bron: Jawahar Swaminathan en MSD -personeel van het European Bioinformatics Institute [Public Domain] via Wikimedia Commons)
Moleculaire structuur van een Hemoolisine (Bron: Jawahar Swaminathan en MSD -personeel van het European Bioinformatics Institute [Public Domain] via Wikimedia Commons) Over het algemeen is Hemoolisine een typisch toxine van de soort van Streptococcus van het darmkanaal. Door de functie kan bacteriën de epitheliale barrière van het darmkanaal breken en zo door de bloedbaan bewegen om andere weefsels te koloniseren.
De meest voorkomende manier waarop hemolisine in de natuur wordt gevonden, is in zijn a-hemolisine-vorm. Dit eiwit is een van de belangrijkste virulentiefactoren van de meeste stammen van Escherichia coli en enkele Clostrides.
De meeste urineweginfecties worden veroorzaakt door stammen van Escherichia coli die a-hemolisine produceren met hemolytische kenmerken.
Hemoolisine- en bacteriocineproductie is gerelateerd aan bacteriestammen met een competentiemechanisme tegen andere soorten en de productie van beide toxines lijkt afhankelijk te zijn van dezelfde genetische determinanten in het bacteriegenoom.
[TOC]
Kenmerken
Hemoolisine wordt gevormd door zeven subeenheden en het gen dat codeert dat het zeven promoters heeft. Deze zeven subeenheden worden ingebracht in het plasmamembraan van de witte cellen en vormen, wanneer ze samen een ionisch kanaal vormen waar de metabolieten van de celinterieur ontsnappen.
Hemoolisine is calciumafhankelijk cytotoxine (Ca+2) extracellulair dat werkt op het plasmamembraan van bloedstroomcellen. De poriën die in het membraan creëren, zijn ook hydrofiel en veroorzaken de binnenkomst van water in het celinterieur, wat lysis kan veroorzaken.
Het kan u van dienst zijn: UNAPORTO: Transport door membranen, kenmerkenHemoolisines zijn eiwitproducten die typerend zijn voor bacteriën van het gramnegatieve type en delen allemaal twee kenmerken:
1- De aanwezigheid van een zeer klein peptide (nonapéptide) gevormd door herhaald glycine en asparaginezuur. Hemoolisine niet-APAPEPTIDE bevinden zich in de buurt van het C-terminale gedeelte van de primaire eiwitstructuur.
2- Alle Hemoolisines worden uitgescheiden door de bacterie op het extracellulaire medium via een ABC-type transporter (van de Engelse ATP-bindende cassette).
Hemoolisines worden meestal gedetecteerd in bacteriestammen door groei in het midden van bloedagar. In de test wordt een hemolytische halo waargenomen, het product van de breuk van de rode bloedcellen in de buurt van de bacteriekolonies.
Jongens
Er zijn verschillende soorten hemoolisines, deze zijn geclassificeerd met een Griekse letter aan het begin van hun naam. De meest bestudeerde en gemeenschappelijke zijn α, β en γ hemoolisines, allemaal geproduceerd door stam Staphylococcus aureus.
Hemoolisine -typen worden geclassificeerd volgens het bereik van cellen die aanvallen en volgens hun primaire eiwitstructuur.
α-hemolisine
Dit eiwit is typerend voor de stammen van Staphylococcus aureus En Escherichia coli; Valt neutrofielen aan, rode bloedcellen, lymfocyten, macrofagen, volwassen en embryonale fibroblasten. Interactie met de polaire koppen van de plasmamembraanlipiden van deze cellen tot het internaliseren van een hydrofobe staart van ongeveer 5 ӑ in het membraan.
β-hemolisine
Gemaakt door Staphylococcus aureus In een mindere verhouding dan α-hemolisine, valt β-hemolisine voornamelijk erytrocyten aan en is het intern in het membraan uitsluitend door rijke domeinen in sfingomyeline van het celmembraan.
γ-hemolisine
Het is ook waargenomen in Staphylococcus aureus. Het is tegelijkertijd geclassificeerd als een hemolytisch eiwit en leukotoxine, omdat het polymorfonucleaire cellen van mensen, monocyten, macrofagen en zelden, zelfs voor de rode bloedcellen beïnvloedt.
Dit type γ-hemolisine is een van de minst gekarakteriseerde, daarom is veel van het werkingsmechanisme onbekend en dit is niet onderzocht In vivo.
Kan u van dienst zijn: osteoblasten: training, kenmerken, functies, pathologieënActiemechanismen
Het werkingsmechanisme dat relatief duidelijk is opgehelderd, is dat van a-hemolisine. Omdat echter allemaal hemolytische eiwitten zijn, wordt gedacht dat de meeste processen gebruikelijk zijn voor alle hemolysines.
Wetenschappers suggereren dat ze voor bacteriën de hemoolisine scheiden naar het milieu dat ze in een micro -omgeving van voedingsstoffen moeten zijn, daarom zou dit een mechanisme zijn dat de cel activeert om witte cellen te vernietigen en hun voedingsstoffen te verkrijgen en hun voedingsstoffen te verkrijgen.
Het mechanisme is in drie stappen beschreven: de unie op het celmembraan, insertie en oligomerisatie.
Membraanunie
Er is gebleken dat hemoolisines zich kunnen aansluiten.
Sommige auteurs suggereren dat de aanwezigheid van receptoren in het membraan niet essentieel is voor de vereniging van hemolysines. In elk geval is het mechanisme van opnieuw aankomende cellulair eiwit nog niet precies bekend.
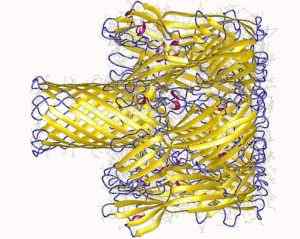 Transmembranale porie gevormd door Staphylococcus Hemoolisine -eiwit (Bron: Depositie Auteurs: Song, L., Hobaugh, m., Shustak, c., Cheley, s., Bayley, H., Gouaux, J.EN.; Visualisatie Auteur: gebruiker: Astrojan [CC door 3.0 (https: // creativeCommons.Org/licenties/door/3.0)] via Wikimedia Commons)
Transmembranale porie gevormd door Staphylococcus Hemoolisine -eiwit (Bron: Depositie Auteurs: Song, L., Hobaugh, m., Shustak, c., Cheley, s., Bayley, H., Gouaux, J.EN.; Visualisatie Auteur: gebruiker: Astrojan [CC door 3.0 (https: // creativeCommons.Org/licenties/door/3.0)] via Wikimedia Commons) De interactie met het membraan vindt plaats in twee stappen:
- Initiële unie (omkeerbaar): wanneer hemolisine bindt aan het membraan calcium junction domeinen. Deze stap vindt plaats op het oppervlak en is zeer gevoelig voor elektrostatische lozingen.
- Onomkeerbare vereniging: boek de aminozuurdomeinen met de lipide -componenten van de buitenste laag van het plasmamembraan van de witte cellen, om fysieke vakbonden te vormen tussen de hydrofobe verbindingen van het membraan.
Kan u van dienst zijn: CelwandInbrengen van toxine in het membraan
Α-hemolisine voegt afval 177 en 411 in de eerste lipidemonolaag. In het extracellulaire medium wordt hemolisine geassocieerd met calciumionen, die hierin een structurele opstelling veroorzaken en bijdragen aan de activering ervan.
Deze insertie consolideert de onomkeerbare unie naar het celmembraan. Zodra de opstelling is opgetreden, wordt hemolisine een integraal eiwit, omdat experimenteel is aangetoond dat de enige manier is om het uit het membraan te extraheren door het gebruik van wasmiddelen zoals Triton X-100.
Oligomerisatie
Wanneer alle Hemoolisine is ingebracht in het plasmamembraan van de witte cel.
Er is waargenomen dat het oligomerisatieproces wordt begunstigd door microdominiums of lipidebalsa's van het celmembraan. Deze regio's bevorderen misschien niet de vereniging van het eiwit, maar zij geven de voorkeur aan de oligomerisatie ervan zodra het is ingevoegd.
Hoe meer hemoolisines aan het membraan binden, hoe groter de hoeveelheid poriën zal worden gevormd. Bovendien kunnen Hemoolisins oligomeriseren met elkaar (aangrenzend) en veel grotere kanalen vormen.
Referenties
- Bakás, l., Ostolaza, h., Vaz, W. L., & Goñi, f. M. (1996). Omkeerbare adsorptie en niet-omkeerbare insertie van Escherichia coli alfa-hemolysine in lipidebilagen. Biophysical Journal, 71 (4), 1869-1876.
- Dalla Serra, m., Coraiola, m., Viero, G., Comai, m., Potrich, c., Ferreras, m.,… & Prévot, g. (2005). Staphylococcus aureus bicomponent γ-hemolysines, HLGA, HLGB en HLGC kunnen gemengd zijn voor het tellen van alle componenten. Journal of Chemical Information and Modellering, 45 (6), 1539-1545.
- Gow, J. NAAR., & Robinson, J. (1969). Eigenschappen van gezuiverde staphylococcale β-hemolysine. Journal of Bacteriology, 97 (3), 1026-1032.
- Ike, en., Hashimoto, h., & Clewell, D. B. (1984). Hemolysine van Streptococcus faecalis Subsecies Zymogenes U draagt bij aan virulentie bij muizen. Infectie en immuniteit, 45 (2), 528-530.
- Remington, J. S., Klein, J. OF., Wilson, c. B., Nizet, v., & Maldonado, en. NAAR. (Eds.)). (1976). Infectieuze woestijnen van de foetus en pasgeboren baby (vol. 4). Philadelphia: Saunders.
- Todd, e. W. (1932). Antigene streptokokkenhemolysine. Journal of Experimental Medicine, 55 (2), 267-280.
- « Pietrain -oorsprong, kenmerken, voedsel, reproductie
- Connivering kleppen kenmerken, histologie, functies »