

Glycolysefuncties, enzymen, fasen, producten, belang
- 3314
- 433
- Aaron Okuneva
De Glycolyse o Glycolyse is de belangrijkste route van het glucosekatabolisme, waarvan de uiteindelijke doel is om energie te verkrijgen in de vorm van ATP en NADH -vermindering van het vermogen, van deze koolhydraat.
Deze route, volledig opgehelderd in de jaren 1930 door Gustav Embden en Otto Meyerhof tijdens het bestuderen van glucoseconsumptie in skeletspiercellen, bestaat uit de volledige oxidatie van genoemde monosaccharide en vertegenwoordigt op zichzelf een anaërobe route voor het verkrijgen van energie.
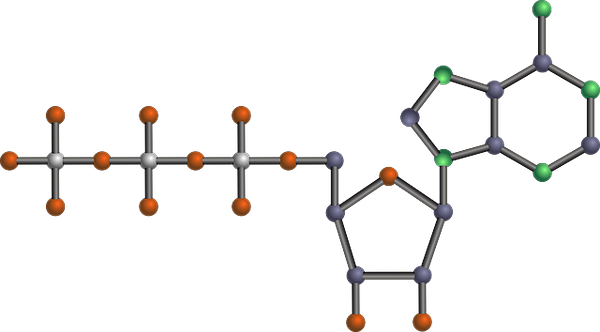
Moleculaire structuur van de ATP, een van de glycolytische producten (samenvatting van de glycolytische route (bron: tekks bij Engelse wikipedia/cc by-sa (https: // creativeCommons.Org/licenties/by-sa/3.0) Via Wikimedia Commons) via Wikimedia Commons) Het is een van de belangrijkste metabole routes, omdat het voorkomt, met zijn verschillen, in alle levende organismen die bestaan, eencellig of multicellulaire, prokaryoten of eukaryoten, en er wordt gedacht dat het een reactieketen is, zeer geconserveerd evolutionair in de natuur in de natuur in de natuur in de natuur.
In feite zijn er enkele organismen en soorten cellen die uitsluitend afhankelijk zijn van deze route naar het leven.
In eerste instantie bestaat glycolyse uit de oxidatie van glucose, van 6 koolstofatomen, tot pyruvaat, dat drie koolstofatomen heeft; Met de gelijktijdige productie van ATP en NADH, nuttig voor cellen vanuit het metabole en synthetische oogpunt.
In die cellen die in staat zijn om de producten te verwerken die zijn verkregen uit glucosekatabolisme, eindigt glycolyse met de productie van koolstofdioxide en water door de Krebs -cyclus en de elektronentransportieketen (aerobe glycolyse).
Tien enzymatische reacties vinden plaats in de loop van de glycolytische route en hoewel de regulatie van deze reacties iets anders kan zijn van de ene soort naar de andere, zijn de regulatiemechanismen ook behoorlijk bewaard gebleven.
[TOC]
Glycolysefuncties
Vanuit het metabolische oogpunt is glucose een van de belangrijkste koolhydraten voor alle levende wezens.
Het is een stabiel en zeer oplosbaar molecuul, dus het kan met relatief gemak worden getransporteerd door het lichaam van een dier of een plant, vanwaar het wordt opgeslagen en/of verkregen tot waar het nodig is als een cellulaire brandstof.
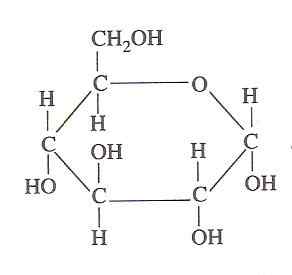 Glucosestructuur (bron: oliva93/cc by-s (https: // creativeCommons.Org/licenties/by-sa/3.0) Via Wikimedia Commons)
Glucosestructuur (bron: oliva93/cc by-s (https: // creativeCommons.Org/licenties/by-sa/3.0) Via Wikimedia Commons) De chemische energie in glucose wordt geëxploiteerd door levende cellen door glycolyse, die bestaat uit een reeks extreem gecontroleerde stappen waardoor de energie die wordt afgegeven uit de oxidatie van deze koolhydraat in meer bruikbare energievormen van energie kan worden "vastgelegd", vandaar het belang ervan kan worden vastgelegd.
Door deze route wordt niet alleen energie (ATP) en het verminderen van kracht (NADH) verkregen, maar het biedt een reeks metabole intermediairs die deel uitmaken van andere routes, ook belangrijk uit de anabole (biosynthetische) en de algemene cellulaire operatie. Hier is een lijst:
- Glucose 6-fosfaat voor de fosfaatroute van Pentosas (PPP, Engels Pantos Mushco -pad))
- Pyruvaat voor melkwegfermentatie
- Pyruvaat voor aminozuursynthese (voornamelijk alanine)
- Pyruvaat voor de tricarbonzuurcyclus
- Fructose 6-fosfaat, glucose 6-fosfaat en dihydroxyacetonfosfaat, die functioneren als "constructieblokken" op andere routes zoals glycogeensynthese, vetzuren, triglyceriden, nucleotiden, aminozuren, enz.
Energie productie
De hoeveelheid ATP geproduceerd door het glycolytische pad.
Als het gaat om aerobe cellen, werkt glycolyse echter ook als een noodzonebron en dient ze als een "voorbereidende stap" voorafgaand aan oxidatieve fosforyleringsreacties die cellen met aerobe metabolisme karakteriseren.
Enzymen die deelnemen aan glycolyse
Glycolyse is alleen mogelijk dankzij de deelname van de 10 enzymen die de reacties katalyseren die deze route karakteriseren. Veel van deze enzymen zijn allosterisch en veranderen vorm of conformatie wanneer ze hun katalytische functies uitoefenen.
Er zijn enzymen die covalente bindingen onder hun substraten breken en vormen en er zijn andere die specifieke cofactoren vereisen om hun functies uit te oefenen, voornamelijk metalen ionen.
Structureel gesproken hebben alle glycolytische enzymen een centrum in wezen gevormd door vellen β parallel omringd door Hellices α en besteld in meer dan één domein. Bovendien worden deze enzymen gekenmerkt omdat hun actieve sites zich meestal in de vakbondslocaties tussen domeinen bevinden.
Het is ook belangrijk om te benadrukken dat de hoofdregulatie van de route door controle (hormonale of metabolieten) van enzymen zoals hexoquinase, fosfofrucerachinase, glyceraldehyde 3-fosfaatdehydrogenase en pyruvaatkinase doorloopt, fosfofrucerachinase, glyceraldehyde.
Kan u van dienst zijn: longparenchym: beschrijving, histologie, ziekten Hoofdreguleringspunten van het glycolytische pad.Org/licenties/by-sa/3.0) Via Wikimedia Commons)
Hoofdreguleringspunten van het glycolytische pad.Org/licenties/by-sa/3.0) Via Wikimedia Commons) 1- Hexoquinase (HK)
De eerste reactie van glycolyse (glucosefosforylering) wordt gekatalyseerd door hexoquinase (HK), waarvan de werkingsmechanisme lijkt te bestaan uit een "geïnduceerde aanpassing" door het substraat, dat de "sluiting" van het enzym rond de ATP en van glucose (HK) lijkt zijn substraten) zodra ze zich hieraan hebben aangesloten.
Afhankelijk van het in aanmerking komende organisme, kunnen er een of meer isoenzymen zijn, waarvan de molecuulgewicht varieert tussen 50 (ongeveer 500 aminozuren) en 100 kDa, omdat ze lijken te zijn gegroepeerd in de vorm van grenzen, waarvan de vorming wordt begunstigd door de Aanwezigheid van glucose, magnesiumionen en ATP.
Hexoquinase heeft een tertiaire structuur bestaande uit alfa- en bètabladen open, hoewel er veel structurele verschillen zijn in deze enzymen.
2- Isomerase-fosfoglucose (PGI)
De gefosforyleerde glucose door hexoquinase wordt isomeriseerd tot 6-fosfaatfructose door middel van isomerase fosfoglucose (PGI), ook bekend als glucose 6-fosfaat isomerase. Het enzym verwijdert of voegt deze niet toe, maar herschikt ze eerder op structureel niveau.
Dit is een actief enzym in zijn dimorvorm (het monomeer weegt min of meer 66 kDa) en is niet alleen betrokken bij glycolyse, maar ook bij gluconeogenese, bij de synthese van koolhydraten in planten, enz.
3- PhospoFRUTEQUINASE (PFK)
De 6-fosfaatfructose is substraat voor het fosfofrucerachinase-enzym, dat in staat is om opnieuw fosforylering te fosforylering met ATP als een fosforylgroepdonor, die 1,6-biffosfaatfructose produceert.
Dit enzym bestaat in bacteriën en zoogdieren als een homotramerisch enzym (samengesteld uit vier identieke subeenheden van 33 kDa elk voor bacteriën en 85 kDa elk bij zoogdieren) en in gisten is het een octameter (samengesteld uit grotere subeenheden tussen 112 en 118 kDa).
Het is een allosterisch enzym, wat betekent dat het positief of negatief wordt gereguleerd door een van zijn producten (ADP) en andere moleculen zoals ATP en citraat.
4- Aldolase
Also known as fructose 1.6-biffosphate Aldolase, aldolase catalyzes the catalytic rupture of fructose 1.6-biphosphate in dihydroxyacetone phosphate and glyceraldehyde 3-phosphate and reaction reaction, that is, the union of both sugars for the formation of the formation of Fructose 1.6-biphosphate.
Met andere woorden, dit enzym snijdt het fructose 1,6-biphosfaat net in het midden, waardoor twee gefosforyleerde verbindingen van 3 koolstofatomen worden vrijgelaten. Aldolase bestaat ook uit 4 identieke subeenheden, elk met een eigen actieve site.
Het bestaan van twee klassen (I en II) van dit enzym is bepaald, die verschillen door het mechanisme van de reactie dat katalyseren en omdat sommige (de eerste) voorkomen in "lagere" bacteriën en eukaryoten, en de anderen (de anderen (de anderen ( de tweede) zijn in bacteriën, protisten en metazoos.
De aldolase van de "superieure" eukaryoten bestaat uit een homotermeter van subeenheden van 40 kDa molecuulgewicht, elk gevormd door een vat samengesteld uit 8 β/α -vellen.
5- Triosa-fosfaat-isomerase (Tim)
De twee Triosas gefosforyleerde kunnen worden verbonden met elkaar dankzij de werking van het isomerase-triosa-fosfaat, waardoor beide suikers kunnen worden gebruikt gedurende glycolyse, waardoor het volledige gebruik van elke glucosemolecule wordt gewaarborgd dat de weg binnenkomt dat de weg binnenkomt.
Dit enzym is beschreven als het "perfecte" enzym, omdat het de reactie van een miljard keer sneller katalyseert dan zonder zijn deelname. De actieve site bevindt zich in het midden van een bèta-vatstructuur, een kenmerk van veel glycolytische enzymen.
Het is een Dimérica -eiwit, gevormd door twee identieke subeenheden van ongeveer 27 kDa, beide met een bolvormige structuur.
6- Glyceraldehyde 3-fosfaatdehydrogenase (GAPDH)
Het 3-fosfaatglyceraldehyde geproduceerd door de werking van het isomerase-triosa-fosfaat dient als een substraat voor de GAPDH, een homoterramerisch enzym (34-38 kDa elke subeenheid) die coöperatief bindt aan een NAD+ molecule in elk van de 4 actieve sites , evenals 2 fosfaat- of sulfaationen.
In deze passage van de route maakt het enzym de fosforylering mogelijk van een van zijn substraten die anorganisch fosfaat gebruikten als een fosforylgroepdonor, met de gelijktijdige reductie van twee NAD+ moleculen en de productie van 1,3-bifospfoglycerato.
7- Fosfoglycerato-kinase (PGK)
Fospoglycerate kinase is verantwoordelijk voor het overbrengen van een van de 1,3-bifosfoglyceraatfosfaatgroepen naar een ADP-molecuul als gevolg van fosforylering op het substraatniveau. Dit enzym maakt gebruik van een mechanisme dat vergelijkbaar is met het gebruik van hexoquinase, omdat het sluit tot contact op zijn substraten en hen beschermt tegen interferentiële watermoleculen.
Kan u van dienst zijn: Aponeurosis: kenmerken, functies, locatieDit enzym heeft, net als anderen die twee of meer substraten gebruiken, een bindende site voor de ADP en een andere voor fosfaatsuiker.
In tegenstelling tot de andere beschreven enzymen, is dit eiwit een 44 kDa -monomeer met een bilobulaire structuur, samengesteld uit twee domeinen van dezelfde grootte verbonden door een smalle "groef".
8-mutase fosfoglyceraat
De 3-fosfoglyceraat ondergaat een verandering van de fosfaatgroep naar koolstof 2, in het midden van het molecuul, dat een strategische instabiliteitsplaats vertegenwoordigt die de daaropvolgende overdracht van de groep naar een ATP-molecuul vergemakkelijkt in de laatste reactie van de route.
Deze Reareglo wordt gekatalyseerd door het enzymfosfoglycerate mutasa, een Dimérica -enzym voor mensen en tetramerisch voor gisten, met een subeenheidgrootte dichtbij 27 kDa.
9- enolase
Enolase katalyseert uitdroging van 2-fosfoglyceraat tot fosfoenolpiruvaat, noodzakelijke stap voor het genereren van ATP in de volgende reactie.
Het is een Dimérica -enzym bestaande uit twee identieke subeenheden van 45 kDa. Het hangt af van magnesiumionen voor stabiliteit en voor de conformationele verandering die nodig is om lid te worden van uw substraat. Het is een van de enzymen uitgedrukt met een grotere overvloed in de cytosol van veel organismen en oefent extra functies uit voor glycolytisch.
10- Quinasa Piruvato
De tweede fosforylering op het substraatniveau dat plaatsvindt in glycolyse wordt gekatalyseerd door het kinasepyruvaat, dat verantwoordelijk is voor de overdracht van de fosforylgroep van het fosfoenolpiruvaat naar de ADP en de productie van pyruvaat.
Dit enzym is complexer dan een van de andere glycolytische enzymen en bij zoogdieren is het een homotramaal enzym (57 kDa/subeenheid). Er zijn, in gewervelde dieren, ten minste 4 isoenzymen: L (in de lever), R (in erytrocyten), M1 (in spier en hersenen) en M2 (foetaal weefsel en volwassen weefsels).
Glycolysefasen (stap voor stap)
De glycolytische route bestaat uit tien opeenvolgende stappen en begint met een glucosemolecuul. Tijdens het proces wordt het glucosemolecuul "geactiveerd" of "voorbereid" met de toevoeging van twee fosfaten, waarbij twee ATP -moleculen worden geïnvesteerd.
Vervolgens wordt het in twee fragmenten "gesneden" en wordt het uiteindelijk een paar keer gemodificeerd, waardoor vier ATP -moleculen worden gesynthetiseerd, dus de netto versterking van de route komt overeen met twee ATP -moleculen.
Uit het bovenstaande kan worden afgeleid dat de route is verdeeld in een energie "investering" -fase, fundamenteel voor de volledige oxidatie van het glucosemolecuul, en een andere fase van energie "gain", waarbij de aanvankelijk gebruikte energie wordt aangevuld en twee ATP -net moleculen.
- Energie -investeringsfase
1- De eerste stap van de glycolytische route bestaat uit de fosforylering van glucose gemedieerd door hexoquinase (HK), waarvoor het enzym een ATP-molecuul gebruikt voor elk glucosemolecuul dat gefosforyleerd is. Het is een onomkeerbare reactie en hangt af van de aanwezigheid van magnesiumionen (mg2+):
Glucose + ATP → Glucose 6-fosfaat + ADP
De aldus geproduceerde aldus geproduceerde 2- 6-fosfaatglucose is geïmantering tot 6-fosfaatfructose dankzij de werking van het isomerase-fosfoglucose-enzym (PGI) (PGI). Dit is een omkeerbare reactie en impliceert geen extra energieverbruik:
Glucose 6-fosfaat → fructose 6-fosfaat
3- Vervolgens impliceert een andere energie-investeringsstap de fosforylering van fructose 6-fosfaat om 1,6-bifosfaatfructose te vormen. Deze reactie wordt gekatalyseerd door het enzym fosfoFRUTEQUINASE-1 (PFK-1). Net als de eerste stap van de route, is het fosfaatgroepdonatormolecuul de ATP en is het ook een onomkeerbare reactie.
Fructose 6-fosfaat + ATP → fructose 1.6-biphosfaat + ADP
4- In deze stap van glycolyse is er de katalytische breuk van fructose 1,6-bifosfaat in dihydroxyacetonfosfaat (DHAP) een ketose, en in glyceraldehyde 3-fosfaat (gap) an aldosa. Deze aldolische condensatie wordt gekatalyseerd door het enzym aldolase en is een omkeerbaar proces.
Fructose 1.6-biffosfaat → dihydroxyaceton fosfaat + glyceraldehyde 3-fosfaat
5- De laatste reactie van de energie-investeringsfase bestaat uit de interconversie van de Triosas Dhap-fosfaat en kloof gekatalyseerd door het triosa-fosfaat isomerase (Tim) enzym, een feit dat geen extra energiebijdrage vereist en dat is ook een omkeerbaar proces.
Dihydroxyaceton fosfaat ↔ glyceraldehyde 3-fosfaat
- Energiewinstfase
6- 3-fosfaat glyceraldehyde wordt "stroomafwaarts" op de glycolytische route gebruikt als een substraat voor een oxidatiereactie en een andere fosforylering, gekatalyseerd door hetzelfde enzym, glyceraldehyde 3-fosfaatdehydrogenase (GAPDH).
Het enzym katalyseert de oxidatie van koolstof C1 van het molecuul tot een carbonzuur en de fosforylering hiervan in dezelfde positie, waardoor 1,3-bifosciceraat wordt geproduceerd. In de loop van de reactie worden 2 NAD+ moleculen verminderd per glucosemolecuul en worden 2 anorganische fosfaatmoleculen gebruikt.
Kan u van dienst zijn: Trachea2Glyceraldehyde 3-fosfaat + 2nad + + 2pi → 2 (1,3-bifospoShoglycerato) + 2nadh + 2H
In aerobe organismen gaat elke NADH die op deze manier wordt geproduceerd door de elektronentransportketen om te dienen als een substraat voor de synthese van 6 ATP -moleculen als gevolg van oxidatieve fosforylering.
7- Dit is de eerste stap van de synthese van ATP in glycolyse en impliceert de werking van de kinasefosfoglycerato (PGK) op 1,3-bifosfoglyceraat, waarbij een fosforylgroep wordt overgebracht (fosforylering op het substraatniveau) van dit molecuul naar een molecuul-ADP, en 2 moleculen van 3-fosfoglyceraat (3pg) voor elk glucosemolecuul.
2 (1.3-bifospospoglycerato) + 2ADP → 2 (3-fosfoglyceraat) + 2Atp
8- The 3-phosphoglycerate serves as a substrate for the enzyme phosphoglycerato mutasa (PGM), which makes it 2-phosphoglycerate by displacement of the fosphoryl group of carbon 3 to carbon 2 by means of a two steps reaction that is reversible and dependent on Magnesiumionen (mg+2).
2 (3-fosfoglyceraat) → 2 (2-fosfoglycerate)
9- Het enzym in een 2-fosfoglyceraat dehydraat en produceert fosfoenolpiruvaat (PEP) door middel van een reactie die de toevoeging van extra energie niet verdient en die tot doel heeft een hoge energieverbinding te produceren, in staat om zijn fosforylgroep in de volgende reactie te schenken in de volgende reactie.
2 (2-fosfoglyceraat) → 2fosfoenolpiruvaat
10- Fosfoenolpiruvaat is een substraat van het pyruvaatkinase (PYK) enzym, dat verantwoordelijk is voor de overdracht van de fosforylgroep in dit molecuul naar een ADP-molecuul, waardoor een andere reactie van fosforylering op het substraatniveau wordt gekatalyseerd.
In de reactie worden 2aTP en 2 pyruvaatmoleculen geproduceerd voor elke glucose en de aanwezigheid van kalium en magnesium in ionische vorm is noodzakelijk.
2fospo elplivato + 2Adp → 2pyruvaat + 2Atp
De netto prestaties van glycolyse bestaat op deze manier uit 2aTP en 2NAD+ voor elk glucosemolecuul dat de route binnengaat.
Als dit cellen zijn met aerobe metabolisme, produceert de totale afbraak van een glucosemolecuul tussen 30 en 32 ATP door de Krebs -cyclus en de elektronentransportketen.
Glycolyseproducten
De algemene reactie van glycolyse is als volgt:
Glucose + 2nad + + 2Adp + 2pi → 2pyruvaat + 2aTp + 2nadh + 2H+
Daarom, als het kort wordt geanalyseerd, kan het ervoor zorgen dat de belangrijkste producten van de glycolytische route pyruvaat, ATP, NADH en H zijn.
De metabolische bestemming van elke reactie -intermediair hangt echter grotendeels af van cellulaire behoeften, en daarom kunnen alle tussenpersonen worden beschouwd als reactieproducten, die ze als volgt kunnen munt:
- Glucose 6-fosfaat
- Fructose 6-fosfaat
- Fructose 1,6-biphosfaat
- Dihydroxyaceton fosfaat en glyceraldehyde 3-fosfaat
- 1.3-bifosfoglycerato
- 3-fosfoglycerate en 2-fosfoglyceraat
- Fosfoenoliruvaat en pyruvaat
Belang
Hoewel glycolyse op zichzelf (zou kunnen praten over anaërobe glycolyse) produceert slechts ongeveer 5% van de ATP die kan worden geëxtraheerd uit aerobe katabolisme van glucose, is deze metabole route om verschillende redenen fundamenteel:
- Het dient als een "snelle" energiebron, vooral in situaties waarin een dier snel een rusttoestand moet verlaten, waarvoor aerobe oxidatieprocessen niet snel genoeg zouden zijn.
- Skeletale "witte" spiervezels in het menselijk lichaam zijn bijvoorbeeld snelle samentrekkingsvezels en zijn afhankelijk van anaërobe glycolyse om te functioneren.
- Wanneer een cel om de een of andere reden glycolytisch nodig heeft.
- Veel cellen zijn afhankelijk van glucose als een energiebron door glycolytische route, inclusief de rode bloedcellen, Het ontbreken van interne organellen en oogcellen (met name die van het hoornvlies) die geen grote mitochondria -dichtheid hebben.
Referenties
- Canback, b., Andersson, s. G. EN., & Kurland, c. G. (2002). De globale fylogenie van glycolytische enzymen. Proceedings of the National Academy of Sciences, 99 (9), 6097-6102.
- Chaudhry R, Varacallo M. Biochemie, glycolyse. [Bijgewerkt 2019 AP 21]. In: StatPearls [internet]. Treasure Island (FL): StatPearls Publishing; 2020 januari-. Beschikbaar vanaf: https: // www.NCBI.NLM.NIH.Gov/books/nbk482303/
- Fothergill-Gilmore, L. NAAR., & Michels, p. NAAR. (1993). Evolutie van glycolyse. Vooruitgang in biofysica en moleculaire biologie, 59 (2), 105-235.
- Kim, J. W., & Dang, c. V. (2005). Veelzijdige rollen van glycolytische enzymen. Trends in Biochemical Sciences, 30 (3), 142-150.
- Kumari, een. (2017). Zoete biochemie: structuren, cycli en routes onthouden door mnemonics. Academische pers.
- Li, x. B., Gu, J. D., & Zhou, Q. H. (2015). Beoordeling van aerobe glycolyse en zijn belangrijkste enzymen-nieuwe doelen voor longkankertherapie. Thoracale kanker, 6 (1), 17-24.
- « Herbivore dierenkenmerken, soorten, voorbeelden
- Tabakmozaïekviruskenmerken, structuur, replicatie »