

Tussenliggende filamentenstructuur, typen, functies
- 2740
- 246
- Pete Heaney V
De tussenliggende filamenten, Ook in de literatuur bekend als "IFS" (uit het Engels Tussenliggende filames), zijn een familie van onoplosbare cytosolische vezelachtige eiwitten die aanwezig zijn in alle meercellige eukaryotische cellen.
Ze maken deel uit van het cytoskelet, een intracellulair filamenteus netwerk dat voornamelijk verantwoordelijk is voor de ondersteuning van de celstructuur en verschillende metabole en fysiologische processen zoals het transport van blaasjes, beweging en celverplaatsing, enz.
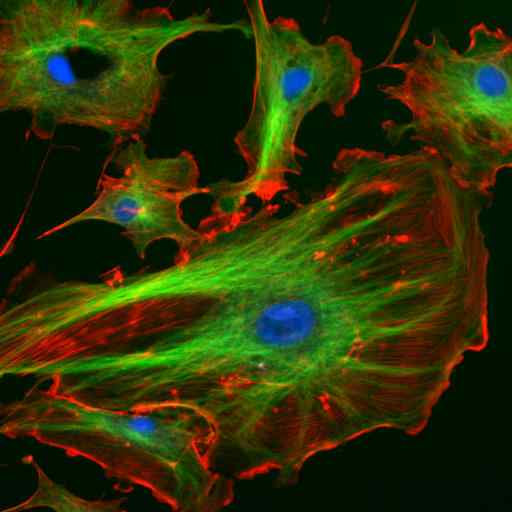
 Microscopie door immunofluorescentie van twee eiwitten van de tussenliggende filamenten van astrocyten (Vimentina en GFAP) (Bron: Gerryshaw [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons)
Microscopie door immunofluorescentie van twee eiwitten van de tussenliggende filamenten van astrocyten (Vimentina en GFAP) (Bron: Gerryshaw [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons) Samen met microtubuli en microfilamenten nemen tussenliggende filamenten deel aan de ruimtelijke organisatie van intracellulaire organellen, in endocytose en exocytose -processen en ook in de processen van celdeling en intercellulaire communicatie.
De eerste tussenliggende filamenten die werden bestudeerd en beschreven waren de keratines, een van de eerste soorten eiwitten waarvan de structuur in de jaren dertig werd geanalyseerd door X -Ray -diffractie.
Het concept van tussenliggende filamenten werd echter geïntroduceerd in de jaren tachtig door Lazarides, die ze beschreven als "mechanische integrators van cellulaire ruimte", gekenmerkt door hun onoplosbaarheid en hun vermogen om opnieuw te bedenken In vitro Na zijn denaturatie.
Veel auteurs worden beschouwd als de "demping" -elementen van stress voor dierlijke cellen, omdat ze flexibelere filamenten zijn dan microtubuli en microfilamenten. Ze worden niet alleen gevonden in het cytoskelet, maar ze maken ook deel uit van de nucleoesqueleto.
In tegenstelling tot de andere vezelige componenten van het cytoskelet, nemen tussenliggende filamenten niet rechtstreeks deel aan celmobiliteitsprocessen, maar werken ze in structureel onderhoud en mechanische weerstand van cellen.
[TOC]
Structuur
Bron: http: // RSB.Info.NIH.Gov/ij/Images/[Public Domain] De tussenliggende filamenten hebben een geschatte diameter van 10 nm, structureel kenmerk waarmee ze werden genoemd, omdat hun grootte tussen de maten is die overeenkomt met de myosine- en actinefilamenten, die respectievelijk ongeveer 25 nm en 7 nm zijn.
Ze verschillen structureel van de andere twee soorten cytoskeletfilamenten, die bolvormige eiwitpolymeren zijn, waarin hun samenstellende eenheden verschillende a-helicooïdale vezelachtige eiwitten zijn van grote lengte die van elkaar zijn gegroepeerd om structuren te vormen in de vorm van touw of kabel.
Alle eiwitten die de tussenliggende filamenten vormen, hebben een vergelijkbare moleculaire organisatie, die bestaat uit een α-helicide of "touw" -domein dat verschillende hoeveelheden "spoelafdeling" van dezelfde grootte heeft.
Dit spiraalvormige domein wordt geflankeerd door een niet-terminale "kop" en een niet-hypoidale "staart" aan het C-terminale uiteinde, en beide variëren zowel in grootte als aminozuursequentie.
Binnen de volgorde van deze twee uitersten zijn de consensusredenen die gebruikelijk zijn voor de 6 soorten bekende tussenliggende filamenten.
In gewervelde dieren heeft het "touw" -domein van het eiwit van de tussenliggende cytosolische filamenten ongeveer 310 aminozuurafval, terwijl de cytosolische eiwitten van de ongewervelde dieren en het nucleaire laminaat ongeveer 350 aminozuren lang hebben.
Kan u van dienst zijn: reptielen: wat zijn, kenmerken, classificatie, reproductieMontage
Tussenliggende filamenten zijn "zelfassemblage" structuren die geen enzymatische activiteit hebben, die hen ook onderscheiden van hun cytoskelet -tegenhangers (microtubuli en microfilamenten).
Deze structuren worden aanvankelijk geassembleerd als tetrameren van filamenteuze eiwitten die hen omvatten onder invloed van alleen monovalente kationen.
Deze tetrameren zijn 62 nm lang en hun monomeren worden zijdelings met elkaar geassocieerd om de "eenheden van lengte" te vormen (UFL, van Engels Eenheid-llege-indieningen), Wat bekend staat als fase 1 van de assemblage, die zeer snel voorkomt.
De UFL's zijn de voorlopers van de lange filamenten en, aangezien de dimeren die hen omvatten elkaar op een antiparalle en verspringende manier binden, hebben deze eenheden een centraal domein met twee flankerende domeinen waardoor fase 2 van de verlenging plaatsvindt, waar de lengteunie plaatsvind van andere UFL's wordt gegeven.
Tijdens wat wordt genoemd als fase 3 van de assemblage, treedt de radiale verdichting van de diameter van de filamenten op, die de volwassen tussenliggende filamenten met meer of minder 10 nm in diameter produceert.
Functie
De functies van de tussenliggende filamenten zijn aanzienlijk afhankelijk van het type cel dat wordt overwogen en, in het geval van dieren (inclusief mensen), wordt hun expressie op een weefselspecifieke manier gereguleerd, dus het hangt ook af van het type weefsel dan onder studie.
Epithelia, spieren, mesenchymale en gliacs en neuronen hebben verschillende soorten filamenten, gespecialiseerd volgens de functie van de cellen waartoe ze behoren.
Onder deze functies zijn de belangrijkste het structurele onderhoud van cellen en weerstand tegen verschillende mechanische stress, omdat deze structuren enige elasticiteit hebben waarmee ze verschillende soorten krachten kunnen dempen die aan cellen worden opgelegd.
Soorten tussenliggende filamenten
Eiwitten die tussenliggende filamenten vormen, behoren tot een grote en heterogene familie van filamenteuze eiwitten die chemisch verschillend zijn, maar worden onderscheiden in zes klassen volgens hun homologie van sequenties (I, II, III, IV, V en VI).
Hoewel het niet erg gebruikelijk is, verschillende soorten cellen, onder zeer specifieke omstandigheden (ontwikkeling, celdransformatie, groei, enz.) Ze kunnen meer dan één klasse van tussenliggende filament die eiwitten vormt samen tot expressie brengen
Klasse I en II van tussenliggende filamenten: zuur- en basis keratines
De keratines vertegenwoordigen de meeste eiwitten van tussenliggende filamenten en bij mensen vertegenwoordigen deze meer dan driekwart van de tussenliggende filamenten.
Ze hebben molecuulgewichten die variëren tussen 40 en 70 kDa en verschillen van andere tussenliggende filamentiwitten vanwege hun hoge gehalte aan glycine en serine.
Kan u van dienst zijn: egels: kenmerken, habitat, reproductie, voedselZe staan bekend als zuur- en basis keratines voor hun iso -elektrische punten, die tussen 4 zijn.9 en 5.4 voor zure keratines en tussen 6.1 en 7.8 voor basics.
In deze twee klassen zijn ongeveer 30 eiwitten beschreven en zijn ze vooral aanwezig in epitheelcellen, waar beide soorten eiwit "co-polimeriseren" en vormen samengestelde filamenten.
Veel van de case i keratines van tussenliggende filamenten zijn te vinden in structuren zoals haar, nagels, hoorns, spikes en klauwen, terwijl klasse II de meest voorkomende in cytosol is.
Klasse III van tussenliggende filamenten: eiwitten van het type demine/vimentine
De watervallen is een zuur eiwit van 53 kDa dat, afhankelijk van de mate van fosforylering, verschillende varianten heeft.
Sommige auteurs hebben ook de filamenten van vallen genoemd als "spierintermediaire filamenten", omdat hun aanwezigheid behoorlijk beperkt is, hoewel in kleine hoeveelheden alle soorten spiercellen.
In myofibrillen wordt de watervallen gevonden in lijn z, dus wordt gedacht dat dit eiwit bijdraagt aan de contractiele functies van spiervezels bij het werken in de vereniging van myofibrilla's en plasmamembraan.
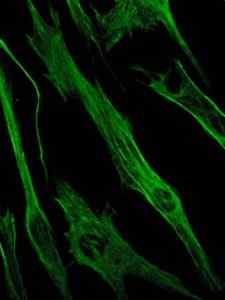 Foto van de kleuring van het vimentine-eiwit, een eiwit van de tussenliggende filamenten van de epitheliale en embryonale cellen (bron: Viktoriia Kosach [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons)
Foto van de kleuring van het vimentine-eiwit, een eiwit van de tussenliggende filamenten van de epitheliale en embryonale cellen (bron: Viktoriia Kosach [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons) Op zijn beurt is vimentine een eiwit aanwezig in mesenchymale cellen. De tussenliggende filamenten gevormd door dit eiwit zijn flexibel en er is vastgesteld dat ze zich verzetten tegen veel van de conformationele veranderingen die tijdens de celcyclus optreden.
Het wordt gevonden in fibroblasten, in gladde spiercellen, in witte bloedcellen en andere cellen van het bloedcirculatiesysteem.
Klasse IV van tussenliggende filamenten: neurofilamenteuze eiwitten
Ook bekend als "neurofilamenten", omvat dit soort tussenliggende filamenten een van de fundamentele structurele elementen van axonen en neuronale dendrieten; Ze worden vaak geassocieerd met de microtubuli die ook deze structuren vormen.
Neurofilamenten van gewervelde dieren zijn geïsoleerd en bepalen dat het een 200, 150 en 68 kDa -eiwittriplet is dat deelneemt aan de assemblage In vitro.
Ze verschillen van andere tussenliggende filamenten waarin ze zijarmen hebben zoals "bijlagen" die worden geprojecteerd vanuit hun periferie en die werken in de interactie tussen naburige filamenten en andere structuren.
Gliacellen produceren een speciaal type tussenliggende filamenten die bekend staan als gliale tussenliggende filamenten, die structureel verschillen van neurofilamenten, omdat ze zijn samengesteld uit een enkel 51 kDa -eiwit en dat verschillende fysicochemische eigenschappen heeft.
Klasse V van tussenliggende filamenten: nucleaire lamina -filamenten
Alle bladen die deel uitmaken van de nucleoesqueleto zijn eigenlijk tussenliggende filamentiwitten. Deze hebben tussen 60 en 75 kDa molecuulgewicht en worden gevonden in de kernen van alle eukaryotische cellen.
Kan u van dienst zijn: Trichuris TrichiuraZe zijn essentieel voor de interne organisatie van de nucleaire regio's en voor veel van de functies van deze essentiële organel voor het bestaan van eukaryoten.
Klasse VI van tussenliggende filamenten: nestines
Dit type tussenliggende filamenten weegt min of meer 200 kDa en bevindt zich voornamelijk in de stamcellen van het centrale zenuwstelsel. Ze worden uitgedrukt tijdens neuronale ontwikkeling.
Gerelateerde pathologieën
Er zijn meerdere ziekten bij mensen die betrekking hebben op tussenliggende filamenten.
Bij sommige soorten kanker, zoals kwaadaardige melanomen of borstcarcinomen, bijvoorbeeld, leiden de co-expressie van tussenliggende vimentine- en keratinefilamenten tot de differentiatie of interconversie van epitheliale en mesenchymale cellen.
Het is experimenteel aangetoond dat dit fenomeen de migrerende en invasieve activiteit van kankercellen verhoogt, wat belangrijke implicaties heeft bij metastatische processen die kenmerkend zijn voor deze aandoening.
Eriksson en medewerkers (2009) beoordelen de verschillende soorten ziekten en hun relatie met specifieke mutaties in de genen die betrokken zijn bij de vorming van de zes soorten tussenliggende filamenten.
Ziekten gerelateerd aan coderende genmutaties voor de twee soorten keratine zijn de bullast epidermolyse, epidermolytische hyperkeratose, cornea dystrofie, keratodermia's en vele andere.
Type III Intermediate filamenten zijn betrokken bij tal van cardiomyopathieën en verschillende spierziekten die voornamelijk verband houden met dystrofieën. Bovendien zijn ze ook verantwoordelijk voor dominante staar en sommige soorten sclerose.
Talrijke neurologische syndromen en aandoeningen zijn gerelateerd aan type IV -filamenten, zoals Parkinson Parkinson. Evenzo zijn genetische defecten in type V- en VI -filamenten verantwoordelijk voor de ontwikkeling van verschillende autosomale ziekten en gerelateerd aan het functioneren van de celkern.
Voorbeeld hiervan zijn onder andere het Hutchinson-Gilford Progeria-syndroom, onder andere Emery-Dreifuss spierdystrofie.
Referenties
- Anderton, B. H. (1981). Tussenliggende Filamles: een familie van homologe structuren. Journal of Muscle Research and Cell Motility, 2(2), 141-166.
- Eriksson, J. EN., Pallari, h., Robert, D., Eriksson, J. EN., Dechat, T., Grijns, B.,… Goldman, r. D. (2009). Introductie van tussenliggende filamoren: van ontdekking tot ziekte. The Journal of Clinical Investigation, 119(7), 1763-1771.
- Fuchs, e., & Weber, K. (1994). Tussenliggende Filamles: structuur, dynamiek, functie en ziekte. Annu. Revisie. Biochem., 63, 345-382.
- Hendrix, m. J. C., Seftor, E. NAAR., Chu, en. W., Trevor, k. T., & Seftor, r. EN. B. (1996). Rol van tussenliggende fillamenten bij migratie, invasie en metastase. Kanker- en metastase -beoordelingen, vijftien(4), 507-525.
- Herrmann, h., & Aebi, u. (2004). Tussenliggende filames: moleculaire structuur, assemblagemechanisme en integratie in functioneel intracellulaire steigers verschillend. Jaaroverzicht van biochemie, 73(1), 749-789.
- Herrmann, h., & Aebi, u. (2016). Tussenliggende Filamles: structuur en montage. Cold Spring Harbor Perspectives in Biology, 8, 1-22.
- McLean, ik., & Lane, B. (negentienvijfennegentig). Tussenliggende filamoren in Diew. Huidige mening in celbiologie, 7(1), 118-125.
- Steinert, P., & Roop, D. (1988). Moleculaire en cellulaire biologie van tussenliggende films. Jaaroverzicht van biochemie, 57(1), 593-625.
- Steinert, P., Jones, J., & Goldman, r. (1984). Tussenliggende filames. The Journal of Cell Biology, 99(1), 1-6.
- « Gastrotrische kenmerken, habitat, reproductie, voedsel
- Soft Moskee -kenmerken, taxonomie, habitat, gebruik »