

Sfingolipiden wat zijn, kenmerken, functies, synthese
- 3758
- 780
- Dr. Rickey Hudson
Wat zijn sphylingolipiden?
De sfingolipiden Ze vertegenwoordigen een van de drie grote lipide families die aanwezig zijn in biologische membranen. Zoals glyceofosfolipiden en sterolen, zijn het amfipatische moleculen met een hydrofiel polair gebied en een hydrofoob apolair gebied.
Ze werden voor het eerst beschreven in 1884 door Johann L. W. Thudichum, die drie sfingolipiden beschreef (sfingomyeline, hersenen en hersenen) die behoren tot de drie verschillende klassen die bekend zijn: fosfosfingolipiden, glycosfingolipiden neutraal en zuur.
In tegenstelling tot glyceofosfolipiden zijn sphyingolipiden niet gebouwd op een 3-fosfaat glycerolmolecuul als het belangrijkste skelet, maar zijn verbindingen afgeleid van SPHINY, een aminoalcohol met een lange koolwaterstofketen bevestigd door een Amida-binding.
ENStructuur van de sfingolipiden
Alle sfingolipiden zijn afgeleid van een L-serine, die is gecondenseerd met een vetzuur met lange keten om de sfingoid-basis te vormen, ook bekend als de Long Chain Base (LCB).
De meest voorkomende basen zijn de sfinganine en sfinxine, die van elkaar verschillen in aanwezigheid van een dubbele binding trans Tussen koolstofatomen 4 en 5 van het vetzuur van de sfinxine.
Koolvesten 1, 2 en 3 van de sfinxine zijn structureel analoog aan glycerol glycerofosfolipiden glycerol. Wanneer Amida -bindingen een vetzuur binden aan koolstof 2 van de sfin.
Vetzuren met lange keten die de hydrofobe gebieden van deze lipiden vormen, kunnen zeer divers zijn. De lengtes variëren van 14 tot 22 koolstofatomen die verschillende verzadigingsgraden kunnen hebben, meestal tussen koolstofatomen 4 en 5.
In posities 4 of 6 kunnen ze hydroxilli -groepen en dubbele bindingen hebben in andere posities of zelfs gevolgen zoals methylgroepen.
Kenmerken
De vetzuurketens die verbonden zijn door amida -bindingen aan de keramides zijn vaak verzadigd en hebben de neiging van grotere lengte te zijn dan die aangetroffen in glycerofosfolipiden, wat cruciaal lijkt te zijn voor de biologische activiteit hiervan.
Een onderscheidend kenmerk van het sfingolipide skelet is dat ze een positieve netto belasting kunnen hebben voor neutrale pH, raar onder lipidemoleculen.
Echter, de PkEen van de aminogroep is laag ten opzichte van een eenvoudig amine, tussen 7 en 8, dus een deel van het molecuul wordt niet geladen tot fysiologische pH, wat de "vrije" beweging hiervan onder bilapas zou kunnen verklaren.
De traditionele classificatie van sfingolipiden komt voort uit de meerdere modificaties die het ceramidemolecuul kan lijden, vooral in termen van de substituties van de polaire hoofdgroepen.
Kan u van dienst zijn: hepadnavirus: kenmerken, morfologie, behandelingFunctie
Sfingolipiden zijn essentieel bij dieren, planten en schimmels, evenals in sommige prokaryotische organismen en virussen.
-Structurele functies
Sfingolipiden moduleren de fysische eigenschappen van membranen, inclusief hun vloeibaarheid, dikte en kromming. Modulair Deze eigenschappen hebben hen ook directe invloed op de ruimtelijke organisatie van membraaneiwitten.
In lipide "vlotten"
In biologische membranen kunnen dynamische micro -domeinen worden gedetecteerd met lagere vloeiendheid gevormd door cholesterol en sfingolipide moleculen genaamd lipidebalsa's.
Deze structuren komen op natuurlijke wijze op en behouden een nauwe relatie met uitgebreide eiwitten, celoppervlakreceptoren en signaaleiwitten, transporters en ander eiwit met glycosylfosfatidylinitol (GPI) ankers (GPI).
-Signaleringsfuncties
Ze hebben functies zoals signaalmoleculen die werken als tweede boodschappers of als uitgescheiden liganden voor celoppervlakreceptoren.
Als secundaire boodschappers kunnen ze deelnemen aan de regulatie van calciumhomeostase, celgroei, tumorogenese en onderdrukking van apoptose. Bovendien hangt de activiteit van veel integrale en perifere membraaneiwitten af van de associatie met sfingolipiden.
Veel intercellulaire en celinteracties met hun omgeving zijn afhankelijk van de blootstelling van de verschillende polaire groepen van de sfingolipiden naar het buitenste gezicht van het plasmamembraan.
De vereniging van glycosfingolipiden en lectina's is cruciaal voor de associatie van myeline met axonen, de hechting van neutrofielen aan het endotheel, enz.
Door -producten van zijn metabolisme
De belangrijkste signaal sfingolipiden zijn de lange ketenbases of de sfinxen en keramides, evenals hun gefosforyleerde derivaten, zoals de 1-fosfaatfaphygosine.
De metabolismeproducten van veel sfingolipiden activeren of remmen meerdere witte downstants (kinase -eiwitten, fosfatose en andere), die cellulair gedrag zo complex regelen als groei, differentiatie en apoptose.
-Als membraanreceptoren
Sommige ziekteverwekkers gebruiken gloysphingolipiden als receptoren om hun ingang tot gastheercellen te bemiddelen of om virulentiefactoren te leveren.
Er is aangetoond dat sphyglyipiden deelnemen aan meerdere celgebeurtenissen zoals secretie, endocytose, chemiotaxis, neurotransmissie, angiogenese en ontsteking.
Ze zijn ook betrokken bij membraanverkeer, dus ze beïnvloeden de internalisatie van receptoren, orde, beweging en fusie van secretoire blaasjes in reactie op verschillende stimuli.
Sfingolipide groepen
Er zijn drie sfingolipide subklassen, allemaal afgeleid van het ceramide en die verschillen van elkaar door polaire groepen, namelijk: sphylingomyelines, glycolipiden en gangliasiden.
Spingomyielinas
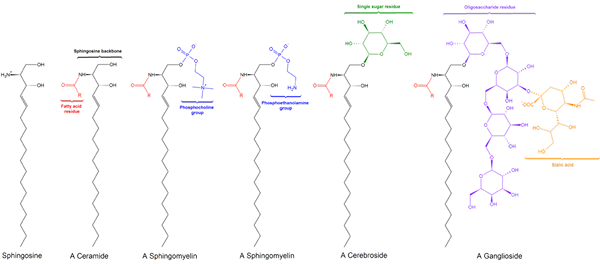
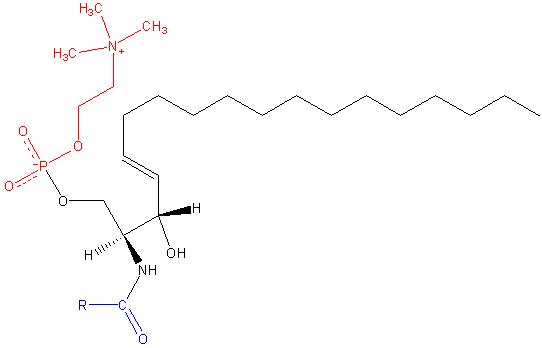 Spingomilein. Zwart: sphinxine. Rood: fosfocoline. Blauw: vetzuur.
Spingomilein. Zwart: sphinxine. Rood: fosfocoline. Blauw: vetzuur. Deze bevatten fosfocoline of fosfoethanolamine als een polaire kopgroep, dus ze worden geclassificeerd als fosfolipiden samen met glyceofosfolipiden. Ze kijken natuurlijk fosfatidylcolines in drie -dimensionale structuur en algemene eigenschappen, omdat ze geen leiding hebben in hun poolhoofden.
Kan u van dienst zijn: enolase: structuur, werkingsmechanisme, functiesZe zijn aanwezig in de plasmamembranen van dierencellen en zijn vooral overvloedig in myeline, een pod die de axonen van sommige neuronen omringt en isoleert.
Glycolipiden of neutrale glycosfingolipiden (zonder belasting)
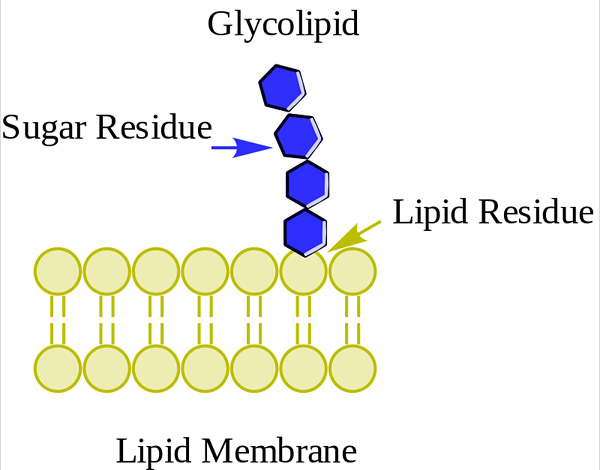 Glucolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons
Glucolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons Ze bevinden zich voornamelijk op de buitenkant van het plasmamembraan en hebben een of meer suikers als een polaire hoofdgroep die direct verenigd is met het koolstofhydroxyl 1 van het keramide -gedeelte. Ze hebben geen fosfaatgroepen. Omdat pH 7 geen belasting heeft, worden ze neutrale glycolipiden genoemd.
Cerebrosiden hebben een enkel suikermolecuul gekoppeld aan ceramide. Degenen die galactose bevatten, bevinden zich in plasmamembranen van niet -nierweefselcellen. Globosiden zijn goustoesphingolipiden met twee of meer suikers, normaal gesproken D-glycose, d-galactose of N-acetyl-d-galactosamine.
Gangliósidos of Glycospingolipiden
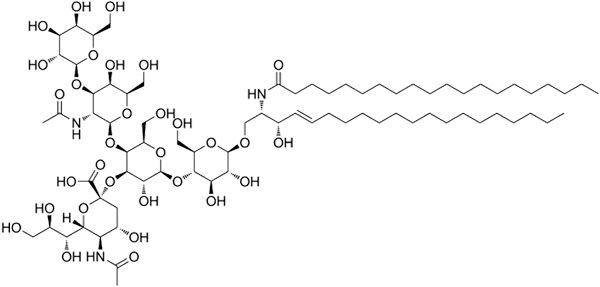 GM1 ganglosidestructuur
GM1 ganglosidestructuur Dit zijn de meest complexe sfingolipiden. Ze hebben oligosachariden als een polaire hoofdgroep en een of meer terminale N-acetylmuraminezuurresiduen, ook wel miralzuur genoemd. Siaminezuur brengt de ganglia de negatieve belasting naar pH 7, die hen onderscheidt van neutrale glycosfingolipiden.
De nomenclatuur van dit soort sfingolipiden hangt af van de hoeveelheid sinaalzuurafval dat aanwezig is in het oligosaccharidegedeelte van de polaire kop.
Synthese
Het lange -keten- of sphiny -basismolecuul wordt gesynthetiseerd in het endoplasmatisch reticulum (ER) en de toevoeging van de polaire groep aan het hoofd van deze lipiden komt later voor in het Golgi -complex. Bij zoogdieren kan enige synthese van sfingolipiden ook voorkomen in mitochondria.
Na het voltooien van hun synthese in het Golgi -complex, worden de sfingolipiden naar andere celcompartimenten getransporteerd via mechanismen gemedieerd door blaasjes.
De biosynthese van de sfingolipiden bestaat uit drie fundamentele gebeurtenissen: de synthese van de lange ketenbases, de biosynthese van de ceramiden door de vereniging van een vetzuur door een Amida -binding, en ten slotte, de vorming van de complexe sfingolipiden door middel van de Unie van polaire groepen in koolstof 1 van de sfingoide -basis.
Naast synthese van novo, Sfingolipiden kunnen zich ook vormen door de vervanging of recycling van de lange kettingbases en keramides, die het sfingolipide pool kunnen voeden.
Ceramide -skeletsynthese
De biosynthese van de keramide, het skelet van de sfingolipiden, begint met de decarboxyleratieve condensatie van een palmitail-CoA-molecuul en een L-serine. De reactie wordt gekatalyseerd door een heterodimeer, pyridoxale fosfaatafhankelijk van een serinepalmitail-transferase (SPT).
Kan u van dienst zijn: microbiota: samenstelling, functies en waar is hetDit enzym wordt geremd door β-phaal-l-alanines en L-cicloserinas. In gisten wordt het gecodeerd door twee genen, terwijl er bij zoogdieren drie genen zijn voor dit enzym. De actieve site bevindt zich in het cytoplasmatische gezicht van het endoplasmatische reticulum.
De rol van dit eerste enzym wordt bewaard in alle bestudeerde organismen. Er zijn echter enkele verschillen tussen taxa die te maken hebben met de subcellulaire locatie van het enzym: bacteriën zijn cytoplasmatisch, gist, planten en dieren bevindt zich in het endoplasmatisch reticulum.
De 3-Zoesfinganine wordt vervolgens verminderd door NADPH-afhankelijke 3-zo-ketosfinganine. Dihydroceramide-synthase (sfinganine N-acil transferase) acetyla vervolgens sfinanine om dihydroceramide te produceren. Het ceramide wordt vervolgens gevormd door de dihydroceramide deaturase/reductase, die een dubbele binding invoegt trans In positie 4-5.
Bij zoogdieren zijn er talloze keramide -isovormen syntasa's, die elk een specifieke vetzuurketen samenvoegen aan de basen van lange ketens. Daarom bieden ceramidensyntheses en andere enzymen, elongasa's, de belangrijkste bron van diversiteit aan vetzuren in sfingolipiden.
Metabolisme
De afbraak van de sfingolipiden wordt uitgevoerd door de glycohydroles en sfingomyelinase -enzymen, die verantwoordelijk zijn voor het verwijderen van de modificaties van de polaire groepen. Aan de andere kant regenereren de ceramidasen de lange kettingbases van de keramides.
De ganglia worden afgebroken door een set lysosomale enzymen die de stap voor stap van de suikereenheden katalyseren, waardoor een ceramide wordt geproduceerd.
Een andere manier van afbraak is de internalisatie van sfingolipiden in endocytische blaasjes die worden teruggestuurd naar het plasmamembraan of naar lysosomen worden getransporteerd waar ze worden afgebroken door specifieke zuurhydrolase.
Niet alle lange ketenbases worden gerecycled, de endoplasmatische behoud. Dit afbraakmechanisme bestaat uit fosforylering in plaats van acilatie van de LCB's, wat leidt tot signaalmoleculen die oplosbare substraten kunnen zijn voor het Liases -enzym.
Regulatie
Het metabolisme van deze lipiden wordt gereguleerd op verschillende niveaus, een daarvan is dat van de enzymen die de leiding geven aan synthese, hun post -translationele modificaties en dezelfde mechanismen van hetzelfde.
Sommige regulatiemechanismen zijn celspecificaties, hetzij om de tijd van celontwikkeling te regelen waarin ze worden geproduceerd of in reactie op specifieke signalen.
Referenties
- Bartke, n., & Hannun, en. (2009). Bioactieve spfingolipiden: metabolisme en functie. Journal of Lipid Research, vijftig, 19.
- Breslow, D. K. (2013). Sfingolipide homeostase in het endoplasmatisch reticulum en verder. Cold Spring Harbor Perspectives in Biology, 5(4), A013326.