

Beperkingsenzymen functies, typen en voorbeelden
- 1364
- 108
- Lonnie Rohan
De Beperkingsenzymen Het zijn endonucleasen die door bepaalde bogen en bacteriën worden gebruikt om de verspreiding van virussen binnen te remmen of te "beperken". Ze zijn vooral gebruikelijk in bacteriën en maken deel uit van hun buitenlandse DNA -afweersysteem dat bekend staat als het restrictiesysteem/modificatiesysteem.
Deze enzymen katalyseren de dubbele band -DNA op specifieke plaatsen, reproduceerbaar en zonder het gebruik van extra energie. De meeste vereisen de aanwezigheid van cofactoren zoals magnesium of andere tweewaardige kationen, hoewel sommige ook ATP- of S-adenosylmetionine nodig hebben.
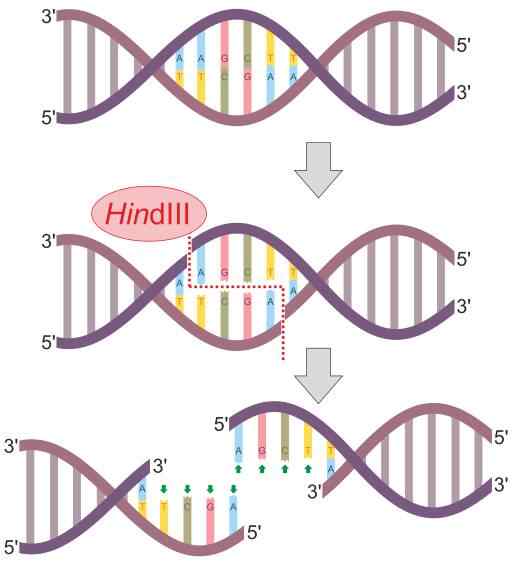
Hindii-restrictie-enzymreactieschema (Bron: Helixitta [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons) De beperking endonucleasen werden ontdekt in 1978 door Daniel Nathans, Arber Werner en Hamilton Smith, die de Nobelprijs voor hun ontdekking ontvingen voor hun ontdekking. Zijn naam komt over het algemeen voort uit het organisme waar ze voor het eerst worden geobserveerd.
Dergelijke enzymen worden veel gebruikt bij de ontwikkeling van DNA -kloneringsmethoden en andere strategieën voor moleculaire biologie en genetische manipulatie. De kenmerken van de herkenning van specifieke sequenties en snijcapaciteit van de sequenties dicht bij de herkenningslocaties maken ze krachtige hulpmiddelen in genetische experimenten.
De fragmenten gegenereerd door de restrictie -enzymen die op een bepaald DNA -molecuul hebben gehandeld, kunnen worden gebruikt om een "kaart" van het oorspronkelijke molecuul te herscheppen door informatie te gebruiken op de locaties waar het enzym het DNA snijdt.
Sommige restrictie -enzymen kunnen dezelfde herkenningslocatie in het DNA hebben, maar ze snijden het niet noodzakelijkerwijs op dezelfde manier. Er zijn dus enzymen die snijwonden maken die romos en enzymen verlaten die het verlaten van samenhangende extreme extremen, die verschillende toepassingen hebben in de moleculaire biologie.
Er zijn momenteel honderden verschillende restrictie -enzymen in de handel verkrijgbaar, aangeboden door verschillende commerciële huizen; Deze enzymen functioneren als "op maat gemaakte" moleculaire schaar voor verschillende doeleinden.
[TOC]
Functie
Beperkingsenzymen vervullen de tegenovergestelde functie van polymerasen, omdat ze worden gehydrolyseerd of de esterbinding verbreekt binnen de fosfodiésterbinding tussen aangrenzende nucleotiden in een nucleotideketen.
In moleculaire biologie en genetische manipulatie zijn ze veel gebruikte hulpmiddelen voor de constructie van expressie en kloneringsvectoren, evenals voor de identificatie van specifieke sequenties. Ze zijn ook nuttig voor de constructie van recombinante genomen en hebben een groot biotechnologisch potentieel.
Recente ontwikkelingen in gentherapie maken actueel gebruik van restrictie -enzymen voor de introductie van genen bepaald in vectoren die voertuigen zijn voor het transport van dergelijke genen naar levende cellen, en dat ze waarschijnlijk het vermogen hebben om zich in het celgenoom in te voegen om permanente veranderingen uit te voeren.
Werkingsmechanisme
Beperkingsenzymen kunnen dubbele band -DNA -snit katalyseren, hoewel sommige in staat zijn om eenvoudige band -DNA -sequenties en zelfs RNA te herkennen. De snede vindt plaats na de herkenning van de sequenties.
Kan u van dienst zijn: glixylaatcyclus: kenmerken, reacties, regulatie, functiesHet werkingsmechanisme bestaat uit de hydrolyse van de fosfodiéster -link tussen een fosfaatgroep en een deoxyribose in het skelet van elk DNA van DNA. Veel van de enzymen zijn in staat om op dezelfde plaats te snijden die ze herkennen, terwijl anderen tussen 5 en 9 basenparen voor of na hetzelfde snijden.
Normaal gesproken snijden deze enzymen aan het 5 'uiteinde van de fosfaatgroep, waardoor DNA -fragmenten aanleiding geven met een 5' fosforyluiteinde en een hydroxyluiteinde 3 'terminal.
Aangezien eiwitten niet in direct contact komen met de DNA -herkenningsplaats, moeten deze opeenvolgende tijden worden getransloceerd totdat de specifieke site is bereikt, misschien door "glijdende" mechanismen op de DNA -streng.
Tijdens de enzymatische snede wordt de fosfodiéster -link van elk van de DNA -strengen geplaatst binnen een van de actieve plaatsen van restrictie -enzymen. Wanneer het enzym de herkennings- en snijlocatie verlaat, doet dit dit door niet -specifieke tijdelijke associaties.
Jongens
Momenteel zijn er vijf soorten restrictie -enzymen bekend. Vervolgens een korte beschrijving van elk:
Type I -beperkingsenzymen
Deze enzymen zijn grote pentamere eiwitten met drie subeenheden, een van beperking, een methylering en een andere voor de herkenning van DNA -sequenties. Deze endonucleasen zijn multifunctionele eiwitten die in staat zijn om beperking en modificatiereacties te katalyseren, ATPASA en ook DNA Topoisomera -activiteit hebben.
De enzymen van dit type waren de eerste endonucleasen die werden ontdekt, ze werden voor het eerst gezuiverd in de jaren zestig en sindsdien zijn ze met grote diepte bestudeerd.
Type I -enzymen worden niet veel gebruikt als een biotechnologisch hulpmiddel, omdat de snijlocatie op een variabele afstand tot 1 kan zijn.000 basenparen met betrekking tot de erkenningssite, waardoor ze onbetrouwbaar zijn in termen van experimentele reproduceerbaarheid.
Type II Beperkingsenzymen
Het zijn enzymen bestaande uit homodimeren of tetrameren die het DNA snijden op locaties die worden gedefinieerd tussen 4 en 8 bp lengte. Deze snijlocaties zijn meestal palindromisch, dat wil zeggen dat ze sequenties herkennen die op dezelfde manier in beide richtingen worden gelezen.
Veel van type II -restrictie -enzymen in bacteriën snijden DNA wanneer ze hun vreemde karakter herkennen, omdat het niet de typische wijzigingen heeft die het eigen DNA zou moeten hebben.
Dit zijn eenvoudiger beperkingsenzymen, omdat ze geen andere cofactor nodig hebben dan magnesium (mg+) om de DNA -sequenties te herkennen en te snijden.
De precisie van type II -restrictie -enzymen bij het herkennen en snijden van eenvoudige sequenties in DNA in precieze posities maakt ze een van de meest gebruikte en onmisbare in de meeste takken van moleculaire biologie.
Kan u van dienst zijn: mutualisme: kenmerken, typen, voorbeeldenBinnen de groep van type II -restrictie -enzymen zijn meerdere subklassen geclassificeerd volgens bepaalde eigenschappen die voor elk uniek zijn. De classificatie van deze enzymen wordt uitgevoerd door letters van het alfabet toe te voegen, van a tot z na de naam van het enzym.
Enkele van de meest bekende subklassen voor hun nut zijn:
Subklasse IIA
Ze zijn verschillende subeenheid Dímeros. Ze herkennen asymmetrische sequenties en worden gebruikt als ideale voorlopers voor het genereren van snijzymen.
Subklasse IIB
Ze zijn samengesteld uit nog een dimeren en snijden het DNA aan beide zijden van de herkenningssequentie. Ze snijden beide DNA -strengen in een basisparen -interval later van de herkenningslocatie.
IIC -subklasse
Enzymen van dit type zijn polypeptiden met divisie- en modificatiefuncties van DNA -strengen. Deze enzymen snijden beide strengen asymmetrisch.
Subklasse IIE
De enzymen van deze subklasse zijn het meest gebruikt in genetische manipulatie. Ze hebben een katalytische plaats en vereisen over het algemeen een alestro -effector. Deze enzymen moeten interageren met twee kopieën van hun herkenningssequentie om een efficiënt snijden te maken. Binnen deze subklasse zijn ECORII- en ECORI -enzymen.
Type III Beperkingsenzymen
Type III Beperking Endonucleasen zijn alleen samengesteld uit twee subeenheden, de ene is verantwoordelijk voor de herkenning en wijziging van DNA, terwijl de andere verantwoordelijk is voor de snede van de volgorde.
Deze enzymen vereisen twee cofactoren voor hun werking: ATP en Magnesium. De restrictie -enzymen van dit type hebben twee asymmetrische herkenningsplaatsen, transloceren het DNA op een ATP -afhankelijke manier en snijden het tussen 20 en 30 bp naast de herkenningsplaats.
Type IV Beperkingsenzymen
Type IV -enzymen zijn gemakkelijk te identificeren, omdat ze DNA met methyleringsmarkeringen snijden, ze bestaan uit verschillende subeenheden die verantwoordelijk zijn voor het herkennen en snijden van de DNA -sequentie. Deze enzymen gebruiken GTP -cofactoren en Divalente Magnesium.
Specifieke snijplaatsen omvatten nucleotideketens met gemethyleerde of hydroxymetyled cytosineresten in een of beide strengen nucleïnezuren.
Type V -restrictie -enzymen
Deze classificatiegroepgroepen scherper-CAS-type enzymen, die specifieke DNA-sequenties van binnenvallende organismen identificeren en snijden. CAS -enzymen gebruiken een RNA -streng gesynthetiseerde gids voor Crisper om binnenvallende organismen te herkennen en aan te vallen.
Enzymen geclassificeerd als type V zijn polypeptiden gestructureerd door type I, II en II -enzymen. Ze kunnen DNA -secties van bijna elk organisme en met een groot bereik van lengte snijden. De flexibiliteit en het gemak van werkgelegenheid maken deze enzymen tot een van de meest gebruikte hulpmiddelen in genetische manipulatie die momenteel samen met Type II -enzymen.
Kan u van dienst zijn: PropionibacteriumVoorbeelden
Beperkingsenzymen zijn gebruikt voor de detectie van DNA -polymorfismen, vooral in populatiegenetica -onderzoeken en evolutionaire studies met behulp van mitochondriaal DNA, om informatie over nucleotidesubstitutie te verkrijgen.
Momenteel hebben de vectoren die worden gebruikt voor de transformatie van bacteriën met verschillende doeleinden multiclonage -locaties waar herkenningsplaatsen voor meerdere beperkingsenzymen worden gevonden.
Onder die enzymen zijn de meest populaire Ecori, II, III, IV en V, verkregen en beschreven voor het eerst van EN. coli; Hindiii, van H. griep en bamhi van B. Amyloliquefaciens.
Referenties
- Bikle, t. NAAR., & Kruger, D. H. (1993). Biologie van DNA -beperking. Microbiologische beoordelingen, 57(2), 434-450.
- Boyaval, p., Moineau, s., Romero, D. NAAR., & Horvath, p. (2007). CRISPR biedt een acqu van virussen in prokaryoten. Wetenschap, 315(Maart), 1709-1713.
- Goodsell, D. (2002). Het moleculaire perspectief: beperking endonucleases. Stamcellen Fundamentals of Cancer Medicine, twintig, 190-191.
- Halford, s. EN. (2001). Hopping, springen en lussen door restrictie -enzymen. Biochemical Society -transacties, 29, 363-373.
- Jeltsch, een. (2003). Onderhoud van soortenidentiteit en controlerende speciatie van bacteriën: een nieuwe functie voor restrictie/modificatiesystemen? Gen, 317, 13-16.
- Krebs, J., Goldstein, E., & Kilpatrick, s. (2018). Lewin's genen xii (12 ed.)). Burlington, Massachusetts: Jones & Bartlett Learning.
- Li, en., Brood, s., Zhang, en., Ren, m., Feng, m., Peng, n.,... zij, q. (2015). Harnasing Type I en Type III CRISPR-CAS-systemen voor genoombewerking. Nucleïnezuren onderzoek, 1-12.
- Loenen, W. NAAR. M., Dryden, D. T. F., Raleight, E. NAAR., & Wilson, G. G. (2013). Type I -beperkingsenzymen en hun familieleden. Nucleïnezuren onderzoek, 1-25.
- Nathans, D., & Smith, h. OF. (1975). Beperking endonucleasen in de analyse en herstructurering van DNA -moleculen. Annu. Revisie. Biochem., 273-293.
- Nei, m., & Tajima, f. (1981). DNA -polymorfisme detecteerbaar door restrictie -endonucleasen. Genetica, 145-163.
- Pingoud, a., Fuxreiter, m., Pingoud, v., & Wende, W. (2005). Cellulaire en moleculaire levenswetenschappen Type II Beperking Endonucleasen: structuur en mechanisme. CML's cellulaire en moleculaire levenswetenschappen, 62, 685-707.
- Roberts, r. (2005). Hoe restrictie -enzymen de werkpaarden van moleculaire biologie werden. PNAS, 102(17), 5905-5908.
- Roberts, r. J., & Murray, K. (1976). Beperking endonucleases. Kritische beoordelingen in biochemie, (November), 123-164.
- Stoddard, B. L. (2005). Homing endonuclease structuur en functie. Driemaandelijkse beoordelingen van biofysica, 1-47.
- Tock, m. R., & Dryden, D. T. F. (2005). De biologie van beperking en anti-herstel. Huidige mening in de microbiologie, 8, 466-472. https: // doi.org/10.1016/j.Mijn B.2005.06.003
- Wilson, g. G., & Murray, n. EN. (1991). Beperkings- en aanpassingssystemen. Annu. Revisie. Genet., 25, 585-627.
- Wu, Z., & Mou, k. (2016). Genomische inzichten in Campylobacter Jejuni virulentie en populatiegenetica. INFEC. Dis. Vertalen. Med., 2(3), 109-19.
- Yuan, r. (1981). Structuur en mechanisme van endonucleasen van multifunctionele restrictie. Annu. Revisie. Biochem., vijftig, 285-315.
- « Generatie van 1914 kenmerken, auteurs en werken
- Wederzijds exclusieve gebeurtenisseneigenschappen en voorbeelden »