

Bilapa lipidische kenmerken, structuur, functies

- 2787
- 675
- Hugo Crooks
De Lipide dubbellaag Het is een dun laminair membraan, bimoleculair, van amfipatische lipiden, dat wil zeggen dat ze een hydrofobe gedeelte en een ander hydrofiel gedeelte bevatten. Het heeft een zeer lage permeabiliteit voor ionen, evenals voor de meeste in water oplosbare moleculen, maar het is zeer permeabel om water te geven.
In waterige oplossingen, polaire lipiden, zoals fosfoglyceride. In deze structuren zijn de hoofden van polaire lipiden, die hydrofiel zijn, extern gericht om in contact te komen met water, terwijl de staarten (hydrofoob) allemaal aan de andere kant zijn gerangschikt.
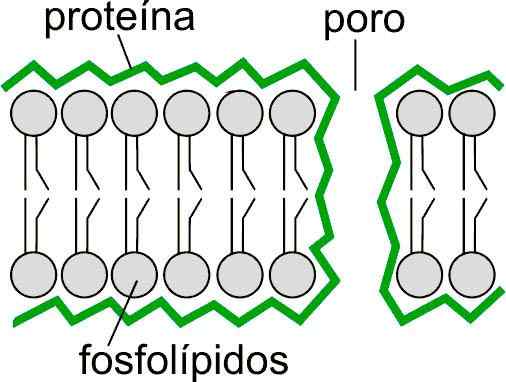
Diagram van mogelijke lipide -opstellingen aan de rand van een porie door een lipide dubbellaag. Genomen en bewerkt uit: mdougm [Public Domain]. Levende wezens hebben celmembranen die voornamelijk zijn gevormd door fosfolipiden en glycolipiden die een lipide dubbellaag vormen. Deze dubbellaag vormt een permeabiliteitsbarrière die het interne gehalte van zouten en celelektrolyten mogelijk maakt. Om dit te kunnen bereiken, hebben ze structuren die ionpompen worden genoemd.
De eerste wetenschappers die het lipide -bilapmodel voor celmembranen voorstelden, waren artsen Evert Gorter en F. Grendel (1925), van de Universiteit van Leiden, Holland, een model dat werd bevestigd in 1950 via elektronische microscopiestudies.
Er zijn verschillende huidige en potentiële toepassingen van lipide bicapas, maar tot op heden is de meest succesvolle commercieel genomen het gebruik van kunstmatige blaasjes (liposomen) in de geneeskunde, voor de toediening van medicijnen aan kankerpatiënten.
[TOC]
Kenmerken
Lipidebilagen zijn laminaire, zeer dunne en fragiele structuren, die enkele biologisch belangrijke kenmerken presenteren, zoals:
Permeabiliteit
Een van de belangrijkste kenmerken van lipide dubbellaag is de selectieve permeabiliteit. Deze membranen zijn inderdaad erg waterdicht voor ionen en de meeste polaire moleculen, water is een belangrijke uitzondering, omdat het gemakkelijk het membraan kan oversteken.
Een voorbeeld van deze selectieve permeabiliteit is natrium en kalium, waarvan de ionen meer dan een miljoen keer langzamer door het membraan gaan dan water. Aan de andere kant kruist de indol, een heterocyclische organische verbinding, het membraan met een snelheid duizend keer hoger dan de tryptofaan, een ander molecuul vergelijkbaar met deze structurele.
Zelfs voordat de dubbele aard van het membraan bekend is, zei de wetenschapper Charles Overton (1901) dat de permeabiliteitscoëfficiënten van de groottemoleculen direct gerelateerd zijn aan de relatieve oplosbaarheid die ze presenteren in organische oplosmiddelen en in water.
Asymmetrie
Elk van de lagen die het membraan vormt, is structureel en functioneel verschillend van de andere. Een functioneel voorbeeld van deze asymmetrie is de natriumkaliumpomp. Deze pomp is aanwezig in het plasmamembraan van de overgrote meerderheid van de cellen van hogere organisme.
Na bom+ - K+ Het is zo georiënteerd dat het na wordt uitgevoerd+ In de cel, terwijl u kionen introduceert+. Bovendien heeft dit transportmiddel energie nodig in de vorm van ATP voor activering en kan alleen worden gebruikt als het zich in de cel bevindt.
De componenten van elke laag zijn ook verschillen.
Kan u van dienst zijn: Crocodylus acutusIn het geval van erytrocyten bevinden zich bijvoorbeeld sphylingomyeline en fosfatidylcolines in de buitenste laag van het membraan, terwijl fosfatidyletalamine en fosfatidylserine intern zijn. Cholesterol is echter samenstellend, beide lagen.
Een van de oorzaken van asymmetrie in de verdeling van fosfolipiden is dat de meeste van deze bestanddelen in de cel worden gesynthetiseerd en daarom in eerste instantie in de binnenste laag worden opgenomen, en daarom zullen sommige van hen naar de buitenlaag migreren met behulp van sommige enzymen die worden genoemd enzymen die worden genoemd enzymen. flipasas.
Vlot
Lipide bilieken zijn geen rigide structuren, maar integendeel, het zijn vloeibare en dynamische structuren, waar lipiden en veel eiwitten moreel moreel moreel moreel zijn.
Lipiden verspreiden zich lateraal in het membraan met een gemiddelde snelheid van 2 µm per seconde. De laterale verplaatsing van eiwitten in dubbellagen kan ondertussen variëren afhankelijk van het type eiwit; Hoewel sommige zo snel zijn als lipiden, blijven anderen vrijwel onbeweeglijk.
De transversale diffusie, ook wel flip-flop genoemd, daarentegen, is veel langzamer voor lipiden en is nooit waargenomen in eiwitten.
Aan de andere kant kan de vloeibaarheid van het membraan variëren afhankelijk van de relatieve volgorde van lipidevetzuren. Wanneer alle vetzuren worden besteld, Bilay.
Deze veranderingen kunnen te wijten zijn aan temperatuurvariaties; De overgang van vaste naar vloeibare toestand vindt abrupt plaats wanneer de temperatuur een drempel overschrijdt die bekend staat als de smelttemperatuur, die afhankelijk is van de lengte van vetzuurketens, evenals de mate van onverzadiging.
Standslipiden van het membraan zijn van verschillende aard en daarom kunnen ze verschillende smelttemperaturen presenteren. Daarom kunnen ze bij verschillende temperaturen naast elkaar bestaan.
Andere mogelijkheden
De lipide dubbellaags hebben, dankzij covalente interacties en aantrekkelijke krachten van van der Waals, de neiging om uitgebreid te zijn, en om in zichzelf te sluiten zodat er geen extreme extreme uitersten zijn. Het vermogen om zelf terug te pakken is ook kenmerkend, omdat een gebrek aan continuïteit niet energetisch gunstig is voor zijn structuur.
Structuur
Er zijn verschillende modellen om de structuur van de lipide dubbellaag te verklaren:
Davson en Danielli Model
In 1935 werd voorgesteld en betoogt dat de membranen een continue koolwaterstoffenfase bevatten, geleverd door samenstellende lipiden van het membraan.
 Davson en Danielli Cell Membrane Model. Genomen en bewerkt uit: Miguelferig [Public Domain].
Davson en Danielli Cell Membrane Model. Genomen en bewerkt uit: Miguelferig [Public Domain]. Eenheidsmembraanmodel
Opgericht door J.D. Robertson, deze hypothese is een aanpassing van het Davson- en Danielli -model. Hij stelde dat het eenheidsmembraan bestond uit een dubbele laag gemengde polaire lipiden.
Deze lipiden waren georiënteerd met de koolwaterstofketens naar binnen en vormden een continue koolwaterstoflaag, terwijl de hydrofiele koppen in de tegenovergestelde richting werden gericht.
Bovendien werd dit unitaire membraan door beide zijden bedekt door een eenvoudige laag eiwitmoleculen gerangschikt uitgebreid.
Bolmodel
Ook bekend als subeenhedenmodel. Volgens dit model zouden de membranen worden gevormd door een mozaïek van repetitieve subeenheden van lipoproteïnen tussen 4,0 en 9,0 nm.
Kan je van dienst zijn: Hammer Head Bat: kenmerken, habitat, reproductie, voedselVloeibaar mozaïek model
Werd voorgesteld door S.J. Zanger en g.L. Nicholson in 1972 en is het meest geaccepteerde model. Volgens het worden membraanfosfolipiden geordend in dubbele lagen, waardoor een matrix van vloeibare kristallen wordt gevormd.
Individuele lipidemoleculen kunnen volgens dit model vrij reizen.
De eiwitten die deel uitmaken van de dubbellager, moeten volgens het model bolvormig zijn. Bovendien zouden sommige eiwitten gedeeltelijk zijn ingebed in de dubbellaag, terwijl anderen hier volledig in zouden zijn ondergedompeld.
De mate van penetratie van bolvormige eiwitten in Bilay.
Samenstelling
Natuurlijke dubbellaags zijn voornamelijk samengesteld uit fosfolipiden. Dit zijn verbindingen afgeleid van glycerol die worden gekenmerkt door een hydrofiele kop en twee hydrofobe staarten.
Wanneer fosfolipiden in contact komen met water kan op verschillende manieren worden georganiseerd. De meest stabiele manier is als dubbellaag met staarten georiënteerd landinwaarts en kopt buiten de dubbellaag.
Glycolipiden maken ook deel uit van de lipide dubbellaagse. Deze verbindingen zijn, zoals de naam al aangeeft, lipiden geassocieerd met suikers, afgeleid in het geval van dieren van een verbinding die bekend staat als sfingocine.
Een ander belangrijk bestanddeel van het membraan is cholesterol, een niet -opscaponificeerbaar lipide. Het is aanwezig zowel in de binnenste als de buitenste laag van de dubbellaag. Het is overvloediger in het plasmamembraan dan in het membraan van de organellen.
De membranen hebben ook veel soorten eiwitten, die van twee soorten kunnen zijn, extrinsiek of intrinsiek. Extrinsieke of perifere eiwitten worden geassocieerd met lakse vakbonden aan het membraan en kunnen ze er gemakkelijk van scheiden.
Intrinsieke of integrale eiwitten zijn sterk geassocieerd met Bilay. Ze vertegenwoordigen ongeveer 70% van de membraaneiwitten. Sommigen van hen hebben een functie van signalen van de buitenkant van de cel en de transmissie ervan binnen.
Andere eiwitten worden geassocieerd met de fusie van twee verschillende dubbellaags. Onder hen zijn degenen die de vereniging van sperma met de eicel toestaan tijdens de bevruchting; Ook die welke virussen toestaan om gastheercellen te penetreren.
Bovendien zijn ionenpompen uitgebreid eiwit dat Bilay kruist.
Functie
De belangrijkste biologische functie van lipide bilay. Zonder deze fysieke afbakening tussen compartimenten zou het leven onmogelijk zijn als we die kennen.
Deze functie is zo belangrijk dat praktisch alle levende wezens een membraan hebben gevormd door een lipide dubbellaag. De uitzondering wordt weergegeven door sommige soorten bogen, waarbij het membraan een lipidemonolaag is.
Lipide dubbellaags nemen deel aan de overdracht van interneuronale zenuwimpuls. Neuronen zijn niet fysiek aan elkaar gekoppeld, maar gescheiden door een korte ruimte genaamd Sinapsis. Om deze interneuronale ruimte te redden, zijn blaasjes geladen met neurotransmitters bij betrokken.
Kan je van dienst zijn: dieren die door de huidmondjes ademenEen andere dubbellaagse functie is om te dienen als een structurele basis of het ondersteunen van skelet, waarvoor sommige transportsystemen sterk verenigd zijn en sommige enzymen.
Organellen met lipide dubbellaagse
In de prokaryoten, de lipidebilay.
-Organellen met twee lipide dubbellaags
Kern
Cellulair organel aanwezig in eukaryotische cellen en dat bevat het grootste deel van het genetische materiaal georganiseerd in chromosomen.
Het nucleaire membraan wordt gevormd door twee lipide bilieken gescheiden door een ruimte die perinucleair wordt genoemd. Beide lagen worden extern en intern nucleair membraan genoemd en worden onderscheiden door de eiwitsamenstelling.
Mitochondria
Organo belast met cellulaire ademhaling, die wordt geleverd door de energie die nodig is voor celactiviteit. Het presenteert een dubbel membraan, het gladde externe en de gevouwen interne vorming laminaire of digitiforme kuif.
De functie van dergelijke plooien is om het interne oppervlak te vergroten, wat de plaats is waar metabole reacties optreden.
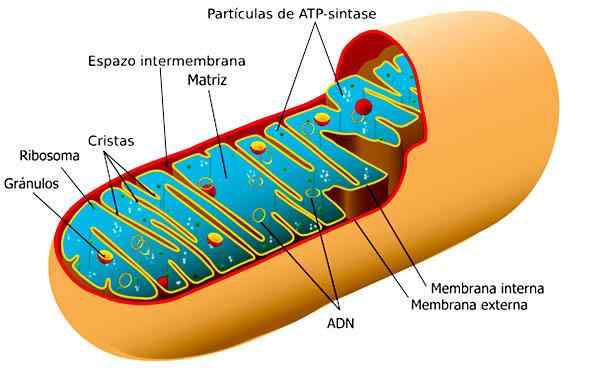 Mitochondria. Genomen en bewerkt uit: ladyofhats [cc0].
Mitochondria. Genomen en bewerkt uit: ladyofhats [cc0]. Chloroplast
Organel aanwezig in bovenste planten en andere fotoautotrofe eukaryotische organismen. Het presenteert twee concentrische lipide bilieken gescheiden door een intermembraneuze ruimte. De buitenste laag is poreuzer dan de interne vanwege de aanwezigheid van eiwitten die porinas worden genoemd.
-Organellen met een lipide dubbellaag
Afgezien van het plasmamembraan, dat in dit artikel op grote schaal is gesproken, hebben andere organellen, zoals het endoplasmatisch reticulum, het Golgi -apparaat en de lysosomen een enkele lipidebilayer.
Endoplasmatisch reticulum (RE)
Geassocieerd cytoplasma membranen complex (re -tostrugged) of niet (soepel RE) met ribosomen, en wie deelneemt aan de synthese van lipiden en fosfolipiden (gladde RE) of peptiden en eiwit (re -ruugous), dankzij ribosomen die aan hun muren zijn bevestigd.
Golgi -apparaat
Complex van gladde wandmembranen die deelnemen aan de opslag, aanpassing en verpakking van eiwitstoffen.
Lysosomen
Vesiculaire organellen die enzymen bevatten die betrokken zijn bij de afbraak van vreemde materialen. Ze degraderen ook niet -noodzakelijke cellulaire componenten en zelfs beschadigde of dode cellen.
Toepassingen
De belangrijkste toepassing van lipide bicapas bevindt zich op het gebied van geneeskunde. Liposomen zijn vesiculaire structuren gescheiden door lipidebilagen. Ze worden kunstmatig gevormd door sonische oscillatie van waterige suspensies van fosfoglyceriden.
Als ionen of moleculen zijn opgenomen in de waterige suspensie, worden sommige van deze elementen in de liposomen opgenomen. Op basis van deze principes zijn geneesmiddelen ingekapseld in oplossing binnen liposomen.
De inhoudelijke liposomen van geneesmiddelen worden geleverd aan de patiënt geïnjecteerd. Eenmaal in het interieur reizen ze door het bloedsysteem, totdat u de doelplaats bereikt. In de bestemming is hun inhoud verbroken en vrijgegeven.
Het gebruik van lipide bilapas zoals biosensoren voor ziektiagnose is ook getest; evenals voor de mogelijke detectie van biologische wapens. Ten slotte is het succesvol getest om de permeabiliteit van het geneesmiddel te bepalen.
Referenties
- OF.S. Andersen, II Koeppe, en. Roger (2007). Bilaagdikte en membraaneiwitfunctie: een energetisch perspectief. Jaaroverzicht van biofysica en biomoleculaire structuur.
- Lipide dubbellaag. In ecured. Hersteld van ecured.com.
- Lipide dubbellaag. In Wikipedia. Hersteld van Wikipedia.borg.
- NAAR. Lehninger (1978). Biochemie. Omega Editions, s.NAAR.
- L. Stryer (1995). Biochemerie. W.H. Freeman and Company, New York.
- R.B. Gennis (1989). Biomembranen. Springer-Verlag.
- M.S. Bretscher (1972). Asymmetrische lipide dubbellaagse structuur voor biologische membranen. Natuur nieuwe biologie.
- « Zenaida Asatische kenmerken, habitat, voedsel
- Harmonia axyridis -kenmerken, levenscyclus, habitat »