

DNA -replicatiemechanismen, in prokaryoten en eukaryoten
- 3251
- 570
- Aaron Okuneva
De Replicatie van DNA (Desoxyribonucleïnezuur) Het bestaat uit het kopiëren van het genoom, dat wil zeggen alle genetische informatie in het DNA van een organisme, om twee identieke kopieën te produceren. Het genoom heeft de nodige informatie om een compleet organisme te bouwen.
Vóór celdeling treedt DNA -replicatie op. Door meiose treden gameten voor seksuele reproductie op. Door mitose treedt celvervanging op (bijvoorbeeld huid en bloed) en ontwikkeling (bijvoorbeeld weefsels en organen).
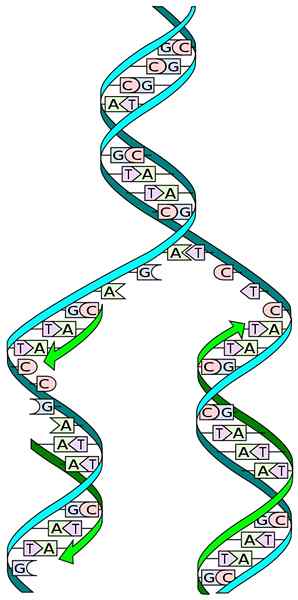
 Bron: I, Madprime [CC0]
Bron: I, Madprime [CC0] De kennis van de DNA -structuur stelt ons in staat om de manier te begrijpen waarop de replicatie plaatsvindt. De structuur van het DNA bestaat uit een dubbele propeller, samengesteld uit twee antipaallale ketens van opeenvolgende nucleotiden, waarvan de stikstofbases specifiek worden aangevuld.
Tijdens de replicatie fungeert elk van de ketens van de dubbele DNA -keten als een vorm voor de biosynthese van een nieuwe keten. De twee nieuw gesynthetiseerde ketens hebben bases die complementair zijn aan de basen van de schimmelketen: adenine (a) met timina (t) en cytosine (c) met guanine (g).
Verschillende enzymen en eiwitten nemen deel aan DNA -replicatie. Bijvoorbeeld het openen van de dubbele DNA-propeller, het DNA openhouden en deoxyribonucleosidos-5'-trifosfaat (DNTP) toevoegen om de nieuwe ketting te vormen.
[TOC]
DNA -replicatie is semi -conservatief
Gebaseerd op de structuur van DNA, stelden Watson en Crick voor dat DNA -replicatie op een semi -conservatieve manier optreedt. Dit werd aangetoond door Meselson en Stahl door het DNA van te markeren Escherichia coli Met de zware isotoop van stikstof, vijftienN, na meerdere generaties het distributiepatroon in een kweekmedium met lichte stikstof, 14N.
Messelson en Stahl ontdekten dat in de eerste generatie de twee dochters -DNA -moleculen hadden, elk molecuul gemarkeerd met een ketting met de zware stikstofisotoop en een andere met de lichte isotoop. In tegenstelling tot het ouderlijke DNA -molecuul, dat de twee ketens had gemarkeerd met de zware isotoop, vijftienN.
In de tweede generatie was 50% van de DNA -moleculen zoals die van de eerste generatie, en de andere 50% had alleen lichte stikstof. De interpretatie van dit resultaat is dat de Double Daughter Propeller een ouderlijke keten heeft (die werkt als een mal) en een nieuwe ketting.
Het semi -conservatieve replicatiemechanisme impliceert de scheiding van DNA -ketens en paring van complementaire basen door middel van opeenvolgende nucleotiden, die twee dochters met dubbele dochters produceren.
Replicatie in batterijen
Start van DNA -replicatie in bacteriën
Bacteriën DNA bestaat uit een circulair chromosoom en heeft slechts één plaats van oorsprong van de replicatie. Van deze site gebeurt de biosynthese van de twee dochters bidirectioneel en vormen twee replicatievork die zich verplaatsen in richtingen die tegen de oorsprong zijn. Uiteindelijk zijn de vorken de replicatie voltooid.
Replicatie begint met de vereniging van DNAA -eiwitten naar de plaats van herkomst. Deze eiwitten vormen op hun beurt een complex. Dan zijn de HU- en IHF -eiwitten binden.
Vervolgens worden DNAC -eiwitten samengevoegd, waardoor de helicasen van DNA zich aansluiten. Deze helpen het DNA af te wikkelen en de waterstofbruggen te breken, gevormd in basenparen. Dus de twee ketens scheiden nog meer, waardoor twee eenvoudige ketens vormen.
Kan u van dienst zijn: wat is chromosomale permutatie?Topoisomerase II, of DNA Girasa, beweegt voor DNA Helicy. DNA -binding van enkele keten (SSB) eiwitten houden de DNA -ketens gescheiden. Zo kan de biosynthese van de dochterketen beginnen.
Biosynthese van DNA -ketens van dochters in bacteriën
Prima -enzym is verantwoordelijk voor het synthetiseren van korte RNA -ketens die primers worden genoemd, die 10 tot 15 nucleotiden lang hebben. DNA-polymerase begint 5'-trifosfaat (DNTP's) toe te voegen aan de 3'-oh van de priming-suiker, waarna de keten met hetzelfde einde blijft groeien.
Omdat DNA -ketens antiparallel zijn, wordt een primer gesynthetiseerd in de geleideketen en veel primers in de vertraagde ketting. Daarom is de biosynthese van de vertraagde keten discontinu. Hoewel DNA -ketens antiparallel zijn, beweegt de replicatievork in één richting.
Het DNA -polymerase is verantwoordelijk voor de vorming van covalente bindingen tussen aangrenzende nucleotiden van de nieuw gesynthetiseerde ketens, in 5'®3 'richting. In EN. coli, Er zijn vijf polymerase DNA: DNA -polymerasen I en III voeren DNA -replicatie uit; en de DNA -polymerasen II, IV en V zijn verantwoordelijk voor het repliceren en repliceren van beschadigd DNA.
Het grootste deel van de replicatie wordt gemaakt door DNA -polymerase III, een holoenzym dat 10 verschillende subeenheden heeft met verschillende functies bij de replicatie van DNA. Alpha -subeenheid is bijvoorbeeld verantwoordelijk voor het maken van banden tussen nucleotiden.
Een enzymcomplex is verantwoordelijk voor de replicatie van DNA in bacteriën
De helicase van DNA en prima is samengevoegd om een complex te vormen dat primosoom wordt genoemd. Dit beweegt langs het DNA en werkt op een gecoördineerde manier om de twee ouderketens te scheiden, waardoor de primers elk bepaald interval op de vertraagde ketting worden gesynthetiseerd.
Primosoom bindt fysiek aan DNA -polymerase III en vormt het replisoom. Twee DNA -polymerasen III zijn verantwoordelijk voor het repliceren van het DNA van de gids en vertraagde ketens. Met betrekking tot DNA -polymerase III vormt de vertraagde keten een lus uit, waardoor de toevoeging van nucleotiden aan deze keten kan plaatsvinden in dezelfde richting van de geleideketen.
De toevoeging van nucleotiden aan de geleideketen is continu. Terwijl het in de vertraging discontinu is. Fragmenten van 150 lengte -nucleotiden worden gevormd, genaamd Okazaki -fragmenten.
De exonuclease -activiteit 5 ' -> 3' van het DNA -polymerase I is verantwoordelijk voor het elimineren van de primers en vulling, het toevoegen van nucleotiden. Een enzymligase verzegelt de openingen tussen fragmenten. Replicatie eindigt wanneer de twee replicatie hoquilla's zich in een voltooiingsreeks bevinden.
Het eiwit bindt aan de beëindigingssequentie, waardoor de beweging van de replicatie -vork wordt gestopt. Topoisomerase II maakt de scheiding van de twee chromosomen mogelijk.
Desoxiribonucleotiden tryposfaten worden gebruikt door DNA -polymerase
Dexinucleoside trypfosfaat (DNTP) bevat drie fosfaatgroepen verenigd met koolstof 5 'van deoxyribosa. De DNTPS (DATP, DTTP, DGTP en DCTP) sluiten zich aan bij de malketen na de AT/GC -regel.
Kan u van dienst zijn: PlesiomorfyHet DNA-polymerase katalyseert de volgende reactie: de hydroxylgroep (-OH) 3 'van het nucleotide van de groeiende keten reageert met het alfa-fosfaat van de inkomende DNTP, waardoor anorganisch pyrofosfaat (PPI) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi) (ppi). PPI -hydrolyse produceert energie voor de vorming van de covalente binding, of fosfodiésterbinding, tussen nucleotiden van de groeiende keten.
Mechanismen die zorgen voor de replicatie -trouw van DNA
Tijdens DNA -replicatie maakt DNA -polymerase III een fout per 100 miljoen nucleotiden. Hoewel de kans op fouten zeer laag is, zijn er mechanismen die zorgen voor betrouwbaarheid in DNA -replicatie. Deze mechanismen zijn:
1) stabiliteit in basisapar. De waterstofbruggenergie tussen AT/GC is groter dan in foutieve basenparen.
2) DNA -polymerase -actieve plaatsstructuur. DNA -polymerase katalyseert bij voorkeur nucleotiden met correcte basen in de tegenovergestelde keten. Een slechte paring van basen veroorzaakt een vervorming van de dubbele DNA -propeller, die voorkomt dat het onjuiste nucleotide de actieve plaats van het enzym bezet.
3) Leestest. DNA -polymerase identificeert opgenomen foutieve nucleotiden en elimineert ze uit de dochterketen. De exonuclease -activiteit van het polymerase -DNA verbreekt de fosfodiésterbindingen tussen nucleotiden aan het 3 'uiteinde van de nieuwe keten.
DNA -replicatie in eukaryoten
In tegenstelling tot de replicatie in prokaryoten, wiens replicatie begint op een enkele site, begint eukaryotische replicatie in meerdere sites van herkomst en de replicatie -vork beweegt bidirectioneel. Vervolgens worden alle replicatievorkjes samengevoegd en vormen ze twee zusterchromatiden die in de centromere zijn samengevoegd.
Eukaryota's hebben veel soorten DNA -polymerase, waarvan de namen Griekse letters gebruiken. DNA -polymerase α vorm een complex met prima. Dit complex synthetiseert korte primers bestaande uit 10 RNA -nucleotiden gevolgd door 20 tot 30 DNA -nucleotiden.
Dan DNA -polymerase ε of δ katalyseert de verlenging van de dochterketen van de primer. DNA -polymerase ε Het is betrokken bij de synthese van de leidende keten, terwijl DNA -polymerase δ Synthetiseer de vertraagde ketting.
DNA -polymerase δ Verleng het linker Okazaki -fragment tot het bereiken van de rechter RNA -primer, waardoor een korte tillen van de primer worden geproduceerd ("korte flap"). In tegenstelling tot prokaryoten, waarbij een polymerase -DNA de primer elimineert, elimineert een endonuclease -enzym in de eukaryoten de RNA -primer.
Vervolgens verzegelt een DNA -ligase de DNA -fragmenten die aangrenzend zijn. De voltooiing van replicatie vindt plaats met de dissociatie van replicatievork -eiwitten.
De DNA -replicatie in eukaryoten en celcyclus
Replicatie in eukaryoten vindt plaats in de S -fase van de celcyclus. Repliceerde DNA -moleculen worden gescheiden in twee dochtercellen tijdens mitose. Fasen G1 en G2 scheiden de S -fase en mitose. De progressie door elke fase van de celcyclus wordt sterk gereguleerd door kinasen, fosfatasen en proteasen.
In de G1 -fase van de celcyclus sluit het oorsprongherkenningscomplex (OCR) zich aan bij de plaats van herkomst. Dit induceert de unie van MCM-helicas en andere eiwitten, zoals Cdc6 en CDT1, om een pre-replicatiecomplex te vormen (Prerc). Las Helicase MCM sluit zich aan bij de gidsketen.
Kan u van dienst zijn: gekoppelde genenIn fase S wordt Prerc een actieve replicatiesite. OCR-, CDC6- en CDT1 -eiwitten worden vrijgegeven en de MCM -helicase beweegt in de richting van 3 'tot 5'. Zodra de replicatie eindigt, wordt dit opnieuw opgestart in de volgende celcyclus.
Replicatie van de uiteinden van chromosomen in eukaryoten
De uiteinden van de chromosomen staan bekend als telomeren, die bestaan uit herhaalde sequenties in combinatie, en uit een 3 'gebied dat uitblinkt, van 12 tot 16 nucleotiden van lengte.
DNA -polymerase kan het 3 'uiteinde van DNA -ketens niet repliceren. Dit komt omdat DNA-polymerase alleen DNA kan synthetiseren in het 5'-3'-adres en alleen reeds bestaande ketens kan verlengen, zonder een primer in deze regio te kunnen synthetiseren. Bijgevolg worden telomeren ingekort in elke replicatieronde.
Telomerase -enzym voorkomt dat het verkorten van telomeren. Telomerase is een enzym dat eiwit- en RNA -subeenheden heeft (derde). De laatste bindt zich aan de herhaalde sequenties van DNA en stelt telomerase in staat zich aan te sluiten bij het 3 'uiteinde van de telomeer.
Een sequentie van RNA achter de vereniging die zich bevindt als een vorm voor de synthese van een sequentie van zes nucleotiden (polymerisatie) aan het einde van de DNA -keten. De verlenging van de telomeer wordt gekatalyseerd door subeenheden van telomerase, genaamd Reverse Telomerase Transcriptase (TERT).
Na polymerisatie vindt translocalisatie plaats, bestaande uit de beweging van telomerase naar een nieuw uiteinde van de DNA -keten, die tot het einde zes andere nucleotiden verbindt.
De functies van andere DNA -polymerasen in de eukaryoten
DNA -polymerase β Het speelt een belangrijke rol bij de eliminatie van onjuiste Bases van DNA, maar het is niet betrokken bij DNA -replicatie.
Velen ontdekte polymerase-DNA behoren tot de polymerasegroep "Translesion-Replicating". Deze polymerasen zijn verantwoordelijk voor het synthetiseren van complementaire ketens in een beschadigd DNA -gebied.
Er zijn verschillende soorten polymerasen "translesion-replicerend". Bijvoorbeeld DNA -polymerase η U kunt repliceren over Timina Dímeros, die worden geproduceerd door UV Light.
DNA -replicatie in archeobacteriën
De replicatie van archeobacteriën DNA is vergelijkbaar met die in eukaryoten. Dit komt door het volgende: 1) eiwitten die deelnemen aan replicatie lijken meer op die van eukaryoten dan aan die van prokaryoten; en 2) Hoewel er slechts één replicatiesite is zoals in prokaryoten, is de volgorde ervan vergelijkbaar met de plaats van eukaryotische oorsprong.
De gelijkenis in de replicatie tussen de bogen en de eukaryoten ondersteunt het idee dat beide groepen fylogenetisch meer gerelateerd zijn aan elkaar dan een van hen met de prokaryoten.
Referenties
- Brooker, r. J. 2018. Genetica -analyse en principes. McGraw-Hill, New York.
- Hartwell, l. H., Goldberg, m. L., Fischer, J. NAAR., Hood, l. 2018. Genetica - van genomen genen. McGraw-Hill, New York.
- Kušić-tišma, j. 2011. Fundamentele aspecten van DNA -replicatie. Intech Open Access, Kroatië.
- Lewis, r., 2015. Menselijke genetica concepten en toepassingen. McGraw-Hill, New York.
- Pierce, B. NAAR. 2005. Genetica - een conceptuele benadering. W. H. Freeman, New York.