

Membraanreceptoren functies, typen, hoe ze werken
- 1465
- 15
- Kurt Aufderhar Jr.
De Membraanreceptoren Ze zijn een type celreceptoren die zich op het oppervlak van het plasmamembraan van de cellen bevinden, waardoor ze chemische stoffen kunnen detecteren die door hun aard het membraan niet kunnen oversteken.
Over het algemeen zijn membraanreceptoren uitgebreide membraaneiwitten die gespecialiseerd zijn in de detectie van chemische signalen zoals peptidehormonen, neurotransmitters en bepaalde trofische factoren; Sommige medicijnen en gifstoffen kunnen ook bij dit soort ontvangers lid worden.
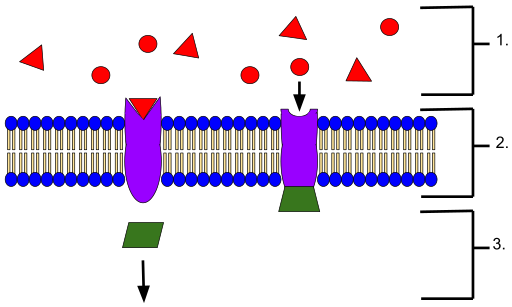
 Representatief schema van een membraanontvanger. De liganden aan de buitenkant van het membraan (1), de membraanligand-receptorinteractie (2) en (3) latere signaalgebeurtenissen (Bron: Wyatt Pyzynski [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons)
Representatief schema van een membraanontvanger. De liganden aan de buitenkant van het membraan (1), de membraanligand-receptorinteractie (2) en (3) latere signaalgebeurtenissen (Bron: Wyatt Pyzynski [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons) Ze zijn geclassificeerd volgens het type intracellulaire waterval waaraan ze zijn gekoppeld en die zijn die het uiteindelijke effect op de overeenkomstige cel bepalen, doelcel of witte cel genoemd.
Daarom zijn drie grote groepen beschreven: die gekoppeld aan ionische kanalen, die gekoppeld aan enzymen en die gekoppeld aan G -eiwit. De unie van liganden aan de ontvangers genereert een conformationele verandering in de ontvanger die een intracellulaire signaalwaterval in de doelcel veroorzaakt.
Signaleringsketens gekoppeld aan membraanreceptoren maken signalen mogelijk om tijdelijke of permanente veranderingen in de doelcel te versterken en te genereren. Deze intracellulaire signalen worden samen "signaaltransductiesysteem" genoemd.
[TOC]
Functie
De functie van membraanreceptoren en andere soorten receptoren in het algemeen is om de communicatie van cellen met elkaar toe te staan, zodat de verschillende organen en systemen van een organisme op een gecoördineerde manier om homeostase te behouden en te reageren op vrijwillige en automatische orders uitgegeven door het zenuwstelsel.
Aldus kan een chemisch signaal dat op het plasmamembraan werkt, een versterkte modificatie van verschillende functies in de biochemische machines van een cel en trigger -multipliciteit van specifieke reacties veroorzaken.
Het kan u van dienst zijn: cadherinas: structuur, kenmerken en functiesVia het signaalamplificatiesysteem is een enkele stimulus (koppelen) in staat om onmiddellijke, indirecte, indirecte veranderingen en langetermijnveranderingen te genereren, waarbij de expressie van sommige genen in de doelcel bijvoorbeeld wordt gemodificeerd, bijvoorbeeld.
Jongens
Cellulaire receptoren worden geclassificeerd, volgens hun locatie, in: membranale receptoren (die worden blootgesteld in het celmembraan) en intracellulaire receptoren (die cytoplasmatisch of nucleair kunnen zijn).
Membraanreceptoren zijn drie typen:
- Gekoppeld aan ionkanalen
- Gekoppeld aan enzymen
- Eiwit G
Membraanreceptoren gekoppeld aan ionische kanalen
Ook wel ionische kanalen genoemd met liganddeur, het zijn samengestelde membraaneiwitten tussen 4 en 6 subeenheden die zodanig worden geassembleerd dat ze een centraal kanaal of porie verlaten, waardoor de ionen van de ene kant naar de andere van het membraan gaan.
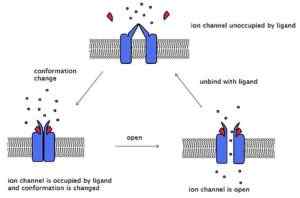 Voorbeeld van de acetylcholinereceptor, een ontvanger gekoppeld aan een ionisch kanaal. De drie conformationele toestanden van dezelfde worden getoond (Bron: Laozhengzz [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons)
Voorbeeld van de acetylcholinereceptor, een ontvanger gekoppeld aan een ionisch kanaal. De drie conformationele toestanden van dezelfde worden getoond (Bron: Laozhengzz [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons) Deze kanalen kruisen het membraan en hebben een extracellulair uiteinde, waar de bindingsplaats zich bevindt, en een ander intracellulair uiteinde dat in sommige kanalen een poortmechanisme presenteert. Bepaalde kanalen hebben een intracellulaire ligandplaats.
Enzym -gebonden membraanreceptoren
Deze receptoren zijn ook transmarket -eiwitten. Ze hebben een extracellulair einde dat de vakbondslocatie voor ligand presenteert en die worden geassocieerd met hun intracellulair einde een enzym dat wordt geactiveerd met de vereniging van het ligand aan de ontvanger.
Membraanreceptoren gekoppeld of eiwit G
GFT -eiwitreceptoren hebben een indirect mechanisme voor de regulatie van intracellulaire functies van doelcellen waarbij transducerende moleculen worden genoemd die fixering of unie -eiwitten van GTP of GTP of eiwitten worden genoemd,.
Kan u van dienst zijn: basaalblad: kenmerken, histologie en functiesAl deze GP -eiwitreceptoren worden gevormd door een membraaneiwit dat zeven keer het membraan kruist en metabotrope receptoren worden genoemd. Honderden receptoren gekoppeld aan verschillende G -eiwitten zijn geïdentificeerd.
Hoe werken ze?
In receptoren gekoppeld aan ionische kanalen genereert de vereniging van het ligand naar de ontvanger een conformationele verandering in de structuur van de ontvanger die een poort kan wijzigen, de wanden van het kanaal kan brengen of verwijderen. Hiermee wijzigen ze de doorgang van ionen van één kant van het membraan aan.
Receptoren gekoppeld aan ionkanalen zijn meestal specifiek voor een iontype, dus receptoren voor K+-kanalen zijn beschreven, van Cl-, van Na+, van Ca ++, enz. Er zijn ook kanalen die twee of meer soorten ionen laten passeren.
De meeste enzymgerelateerde receptoren zijn geassocieerd met kinase -eiwitten, vooral met het kinasetyrosine -enzym. Deze kinasen worden geactiveerd wanneer de receptorbinding op zijn extracellulaire vakbondslocatie. Quinaas fosforyleren specifieke eiwitten in de doelcel, die de functie van hetzelfde wijzigt.
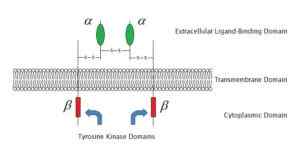 Voorbeeld van membraanreceptor gekoppeld aan het enzym tyrosinekinase (Bron: Laozhengzz [Public Domain] via Wikimedia Commons)
Voorbeeld van membraanreceptor gekoppeld aan het enzym tyrosinekinase (Bron: Laozhengzz [Public Domain] via Wikimedia Commons) Eiwit -gebogen receptoren activeren.
Er zijn twee soorten G -eiwitten die heterotrimische G -eiwitten en monomere G -eiwitten zijn. Beide zijn inactief gekoppeld aan het BBP, maar wanneer het ligand naar de ontvanger wordt vervangen door het BBP door GTP en het G -eiwit wordt geactiveerd.
In heterotrimische G -eiwitten dissocieert de α -subeenheid gekoppeld aan de GTP uit het ßy -complex, waardoor het eiwit G geactiveerd blijft. Zowel de α -subeenheid gekoppeld aan de GTP, en de vrije ßy kan het antwoord bemiddelen.
Kan je van dienst zijn: trofoblast Schema van een GF -eiwit gekoppelde ontvanger (Bron: Bensacount bij Engelse Wikipedia [Public Domain] via Wikimedia Commons)
Schema van een GF -eiwit gekoppelde ontvanger (Bron: Bensacount bij Engelse Wikipedia [Public Domain] via Wikimedia Commons) Monomere G -eiwitten of kleine G -eiwitten ontvangen ook de naam van RAS -eiwitten omdat ze voor het eerst worden beschreven in een virus dat sarcommon tumoren bij ratten produceert.
Wanneer geactiveerd, stimuleren ze mechanismen voornamelijk gerelateerd aan vesiculair verkeer en cytoskeletfuncties (modificatie, remodellering, transport, enz.)).
Voorbeelden
De acetylcholinereceptor, gekoppeld aan een natriumkanaal dat opent bij het verbinden van acetylcholine en depolarisatie van de doelcel genereert, is een goed voorbeeld van membraanreceptoren gekoppeld aan ionische kanalen. Bovendien zijn er drie soorten glutamaatreceptoren die ionotrope receptoren zijn.
Glutamaat is een van de belangrijkste excitatieve neurotransmitters van het zenuwstelsel. De drie soorten ionotrope receptoren zijn: de NMDA (N-methyl-D-spartaat), AMPA (a-amino-3-hydroxy-5-methyl-4-4-isoxazol-propionaat) en kainato) en kainato (zure kainico).
Hun namen zijn afgeleid van de agonisten die ze activeren en deze drie soorten kanalen zijn voorbeelden van unieke spannende kanalen, omdat ze de doorgang van natrium en kalium en in sommige gevallen van kleine hoeveelheden calcium mogelijk maken.
Voorbeelden van enzym -gekoppelde receptoren zijn insulinereceptor, de familie van TRK -receptoren of neurotrofinereceptoren en receptoren voor sommige groeifactoren.
Onder de belangrijkste eiwit G-receptoren zijn de acetylcholinemuscarine-receptoren, β-adrenerge receptoren, reuksysteemreceptoren, metabotrope glutamaatreceptoren, receptoren voor veel peptide-hormonen en rodopsineceptoren van het retino-systeem.
Referenties
- Afdeling Biochemie en Molecular Biophysics Thomas Jesell, Siegelbaum, S., & Hudspeth, een. J. (2000). Principes van neurale wetenschap (Vol. 4, pp. 1227-1246). EN. R. Kandel, J. H. Schwartz, & T. M. Jesell (eds.)). New York: McGraw-Hill.
- Hulme, e. C., Birdsall, n. J. M., & Buckley, n. J. (1990). Muscarinische ontvanger subtypen. Jaaroverzicht van farmacologie en toxicologie, 30(1), 633-673.
- Cull-Candy, S. G., & Leszkiewicz, D. N. (2004). Rol van verschillende NMDA -ontvangersubtypen bij centrale synapsen. Sci. Stke, 2004(255), RE16-RE16.
- William, f. G., & Ganong, m. D. (2005). Overzicht van medische fysiologie. Gedrukt in de Verenigde Staten van Amerika, zevende editie, PP-781.
- Beer, m. F., Connors, B. W., & Paradiso, m. NAAR. (Eds.)). (2007). Neurowetenschap (Vol. 2). Lippinott Williams & Wilkins.
- « Achlostoma caninum -kenmerken, morfologie, symptomen
- Canelo -kenmerken, habitat, eigenschappen, teelt »