

Microfilamentenkenmerken, structuur, functies, pathologie
- 3656
- 1086
- Nathan Wiegand
De microfilamenten o Actinefilamenten, zijn een van de drie fundamentele componenten van eukaryotische celcytoskelet.
In eukaryoten zijn de genen die coderen voor actine -microfilamenten sterk bewaard in alle organismen, dus ze worden vaak gebruikt als moleculaire markers voor verschillende onderzoeken.
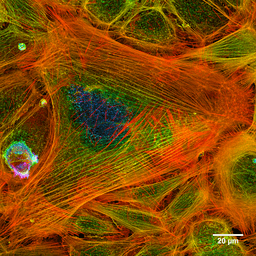
Foto van de actinefilamenten van een geverfde cel (Bron: Howard Vindin [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons) Microfilamenten worden over het hele cytosol verdeeld, maar ze zijn vooral overvloedig in het onderliggende gebied naar het plasmamembraan, waar ze een complex netwerk vormen en worden geassocieerd met andere speciale eiwitten om het cytoskelet te vormen.
Microfilamentnetwerken in het cytoplasma van zoogdiercellen worden gecodeerd door twee van de zes genen die worden beschreven voor actine, die betrokken zijn bij de dynamiek van microfilamenten en die zelfs erg belangrijk zijn tijdens de differentiatie van de stamcellen.
Veel auteurs zijn het erover eens dat microfilamenten de meest diverse, veelzijdige en belangrijke eiwitten van het cytoskelet van de meeste eukaryotische cellen zijn, en het is belangrijk om te onthouden dat deze niet worden gevonden in prokaryotische micro -organismen.
In dit type cellen zijn er echter homologe filamenten voor microfilamenten, maar die worden gevormd door een ander eiwit: MREB -eiwit.
Momenteel wordt gedacht dat het gen dat voor dit eiwit codeert een mogelijk voorouderlijk gen is voor eukaryotisch actine. De aminozuursequentiehomologie die MREB -eiwit vormt, is echter slechts 15% ten opzichte van de actine -sequentie.
Net als een fundamenteel onderdeel van het cytoskelet, kan elk fenotypisch defect zowel in microtubuli als in tussenliggende filamenten en actinemicrofilamenten (cytoskelet) verschillende cel- en systemische pathologieën veroorzaken.
[TOC]
Kenmerken en structuur
Microfilamenten bestaan uit eiwitmonomeren van de actinefamilie, die extreem overvloedige contractiele eiwitten in eukaryotische cellen zijn, omdat ze ook deelnemen aan spiercontractie.
Deze filamenten hebben een diameter tussen 5 en 7 nm, dus ze staan ook bekend als dunne filamenten en zijn samengesteld uit twee vormen van actine: de bolvormige vorm (actin g) en de filamenteuze vorm (actine F).
Eiwitten die deelnemen aan het cytoskelet staan bekend als y- en β -actine, terwijl degenen die deelnemen aan contractie meestal actines α zijn.
Kan u van dienst zijn: cytoplasma: functies, onderdelen en kenmerkenHet aandeel van bolvormige actine en filamenteuze actine in het cytosol hangt af van cellulaire behoeften, omdat microfilamenten zeer variabel zijn en veelzijdige structuren zijn, die voortdurend groei en verkorting zijn als gevolg van polymerisatie en depolymeterisatie.
Actin G is een klein bolvormig eiwit, bestaande uit bijna 400 aminozuren en ongeveer 43 kDa molecuulgewicht.
Actineer monomeren waaruit de microfilamenten bestaat, worden besteld in de vorm van een spiraalvormige streng, omdat elk een torsie lijdt wanneer ze worden geassocieerd met het volgende.
Actin G wordt geassocieerd met een Ca2+ -molecuul en een andere ATP, die zijn bolvormige vorm stabiliseren; Terwijl actine F wordt verkregen na de hydrolyse van het terminale fosfaat van het ATP -molecuul in actine, wat bijdraagt aan polymerisatie.
Organisatie
Actinefilamenten kunnen worden georganiseerd in de vorm van "balken" of "netwerken" die verschillende functies in de cellen hebben. De stralen vormen parallelle structuren gekoppeld door vrij rigide kruisbruggen.
Netwerken daarentegen zijn comfortabelere structuren, zoals drie -dimensionale mazen met de eigenschappen van semi -solid gels.
Er zijn veel eiwitten die worden geassocieerd met actine- of microfilamentfilamenten en die bekend staan als ABP (uit het Engels Actinebindende eiwitten), die hiervoor specifieke sites hebben.
Veel van deze eiwitten stellen microfilamenten in staat om te interageren met de andere twee componenten van het cytoskelet: microtubuli en tussenliggende filamenten, evenals met de andere componenten van het binnenste gezicht van het plasmamembraan.
Onder andere eiwitten die interageren met microfilamenten zijn nucleaire vellen en spectrine (in rode bloedcellen).
Hoe worden actinefilamenten gevormd?
Omdat bolvormige actinemonomeren altijd op dezelfde manier meedoen, georiënteerd in dezelfde richting, hebben microfilamenten een gedefinieerde polariteit, met twee uitersten: één "meer" en één "minder".
De polariteit van deze filamenten is erg belangrijk, omdat ze aanzienlijk sneller worden vanwege hun positieve einde, waar de nieuwe actine -monomeren worden toegevoegd.
 Grafische weergave van actine -microfilamenten (Bron: Derivative Work: Retama (Talk) Thin_Filament_Formation.SVG: Mikael Häggström [Public Domain] via Wikimedia Commons)
Grafische weergave van actine -microfilamenten (Bron: Derivative Work: Retama (Talk) Thin_Filament_Formation.SVG: Mikael Häggström [Public Domain] via Wikimedia Commons) Het eerste dat plaatsvindt tijdens de polymerisatie van actinefilamenten is een proces dat bekend staat als "nucleatie", dat bestaat uit de associatie van drie eiwitmonomeren.
Kan u van dienst zijn: plasomolyseAan deze trimmer worden nieuwe monomeren aan beide uiteinden toegevoegd, zodat de gloeidij groeit. Actin G-monomeren kunnen ATP hydrolyseren met elke unie, die implicaties heeft in de snelheid van polymerisatie, omdat actine-ATP-delen meer moeite dissociëren dan actine-ADP.
De ATP is niet nodig voor polymerisatie en de betonfunctie van zijn hydrolyse is nog niet verduidelijkt.
Sommige auteurs zijn van mening dat, aangezien actinepolymerisatie -evenementen snel zijn.
Regulatie
Zowel de polymerisatie van actinefilamenten als de depolimerisatie ervan zijn processen die sterk worden gereguleerd door een reeks specifieke eiwitten, die verantwoordelijk zijn voor de remodellering van de filamenten.
Voorbeeld van de eiwitten die depolymeren reguleren. Een ander eiwit, profylaine, heeft een tegenovergestelde functie, omdat het de associatie van monomeren stimuleert (door de uitwisseling van ADP voor ATP te stimuleren).
Functie
Microfilamenten interageren met myosine -filamenten die worden geassocieerd met transmembraaneiwitten die een domein hebben in cytosol en een andere in het buitenland -cel, zodat ze deelnemen aan celmobiliteitsprocessen.
Deze microfilamenten geassocieerd met het plasmamembraan bemiddelen verschillende cellulaire responsen op verschillende soorten stimuli. Celadhesie van epitheliale weefsels wordt bijvoorbeeld geleid door transmembraaneiwitten die bekend staan als Cadherinas, die interageren met microfilamenten om responsfactoren te werven.
Actinefilamenten interageren met tussenliggende filamenten om extracellulaire stimuli te laten overbrengen naar belangrijke plaatsen zoals ribosomen en chromosomen in de kern.
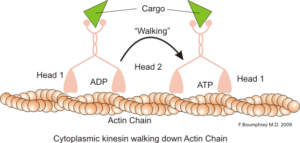 Representatie van de intracellulaire motorfunctie van actinemicrofilamenten (Bron: Boumphreyfr [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons)
Representatie van de intracellulaire motorfunctie van actinemicrofilamenten (Bron: Boumphreyfr [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons) Een klassieke en zeer bestudeerde functie van microfilamenten is het vermogen om "bruggen", "rails" of "snelwegen" te vormen voor de beweging van myosinemotor eiwit I, die in staat is om transportblaasjes van de organellen naar het membraanplasma op het secretory te laden paden.
Microfilamenten interageren ook met myosine II om de contractiele ring te vestigen die wordt gevormd tijdens cytokinese, precies tijdens de laatste fase van de celdeling waarin de cytosol wordt gescheiden van de stam- en dochtercellen.
Kan u van dienst zijn: polymorfonucleaire leukocytenOver het algemeen moduleren actine -vormige microfilamenten de verdeling van sommige organellen zoals het Golgi -complex, het endoplasmatisch reticulum en mitochondria. Bovendien nemen ze ook deel aan de ruimtelijke positionering van de RNM's zodat ze door de ribosomen worden gelezen.
De hele set van de mobiele telefoon van microfilamenten, vooral die die nauw verwant zijn met het plasmamembraan, neemt deel aan de vorming van golvende celmembranen van cellen die een constante actieve beweging hebben.
Ze nemen ook deel aan de vorming van microvings en andere gemeenschappelijke uitsteeksels op het oppervlak van veel cellen.
Voorbeeld van functies in de lever
Microfilamenten nemen deel aan het galafscheidingsproces in hepatocyten (levercellen) en ook aan peristaltische bewegingen (gecoördineerde contractie) van leverkaniculi.
Ze dragen bij aan de differentiatie van plasmamembraandomeinen dankzij hun associatie met verschillende cytosolische elementen en de controle die ze uitoefenen bij de topografie van deze intracellulaire elementen.
Gerelateerde pathologieën
Er zijn weinig ziekten geassocieerd met primaire defecten in de structuur of met eiwitten en regulerende enzymen in de synthese van microfilamenten, hoewel deze direct bij een groot aantal functies betrokken zijn.
De lage index van ziekten en misvormingen in de primaire structuur van microfilamenten is te wijten aan het feit dat er in het algemeen meerdere coderende genen van zowel actine als zijn regulerende eiwitten zijn, een fenomeen dat bekend staat als "genetische redundantie".
Een van de meest bestudeerde pathologieën is de vitrificatie van eicellen op hun cytoskelet, waarbij een onderbreking wordt waargenomen in het corticale microfilamentnetwerk, evenals een depolimerisatie en een desorganisatie van de microtubuli van de mitotische spil.
In het algemeen veroorzaakt deze vitrificatie chromosomale dispersie, omdat het leidt tot een meningsverschil bij de verdichting van de gehele chromatine.
De cellen met een grotere organisatie en het aandeel van microfilamenten in hun cytoskelet zijn gestreepte spiercellen, daarom zijn de meeste pathologieën geassocieerd met een storing van het contractiele apparaat.
Defecte of atypische microfilamenten zijn ook geassocieerd met botziekte die bekend staat als Paget -ziekte.
Referenties
- Aguilar-Cuenca, R., Llorente-González, C., Vicente, c., & Vicente-Manzanares, m. (2017). Microfilament-gecoördineerde adhesiedynamiek stimuleert enkele celmigratie en vormt wisselen. F1000resarch, 6.
- Twee remedies, c. G., Chhabra, D., Kekic, m., Vinger, ik. V., Tsubakihara, m., Berry, D. NAAR., & Nosworthy, n. J. (2003). Bindende eiwitten actine: regulatie van cytoskeletmicrofilamenten. Fysiologische beoordelingen, 83(2), 433-473.
- Guo, h., Fauci, l., Shelley, m., & Kanso, en. (2018). Bistabiliteit in de synchronisatie van geactiveerde microfilamenten. Journal of Fluid Mechanics, 836, 304-323.
- Launch., Langer, r., & Vacanti, j. P. (Eds.)). (2011). Principes of Tissue Engineering. Academische pers.
- Robbins, J. (2017). Ziekten van het cytoskelet: de deminopathieën. In cardioskeletale myopathieën bij kinderen en jullie volwassenen (PP. 173-192). Academische pers.
- « Grizzly Bear -kenmerken, habitat, voedsel, gedrag
- Cistus clusii -kenmerken, taxonomie, habitat, distributie, gebruik »