

Humorale immuniteitstheorie, effectormechanismen, voorbeelden
- 4091
- 347
- Irving McClure I
De humorale immuniteit, Ook bekend als de antilichaam -gemedieerde immuunrespons, is het een van de belangrijkste afweermechanismen van organismen tegen invasie uit extracellulaire micro -organismen of toxines.
In het bijzonder verwijst humorale immuniteit naar immuniteit gemedieerd door bloedfactoren, die serumeiwitten zijn die bekend staan als "antilichamen" die werken in reacties op infecties en die specifiek worden geproduceerd in reactie op de aanwezigheid van "antigenen".
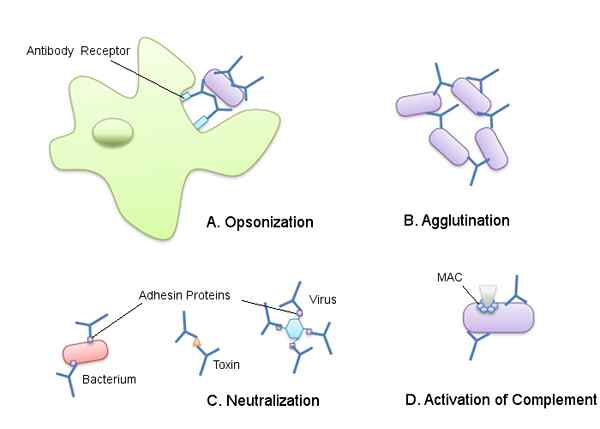 Sommige van de effecten van de antilichamen geproduceerd tijdens de humorale immuunrespons (Bron: Becky Boone [CC BY-SA 2.0 (https: // creativeCommons.Org/licenties/by-sa/2.0)] via Wikimedia Commons)
Sommige van de effecten van de antilichamen geproduceerd tijdens de humorale immuunrespons (Bron: Becky Boone [CC BY-SA 2.0 (https: // creativeCommons.Org/licenties/by-sa/2.0)] via Wikimedia Commons) Het immuunsysteem van een zoogdier kan worden onderverdeeld in aangeboren immuunsysteem en adaptief immuunsysteem. Het aangeboren immuunsysteem bestaat uit de verschillende elementen die fungeren als fysische en chemische barrières tegen de binnenkomst van middelen die aan het lichaam binnenvallen.
Onder dergelijke barrières zijn de epithelia en sommige van de door hen geproduceerde stoffen; Sommige soorten specifieke cellen nemen ook deel die samen het eerste lichaamsdefensiesysteem vertegenwoordigen.
Het adaptieve of specifieke immuunsysteem is een beetje complexer en "geëvolueerd", omdat het wordt geactiveerd als reactie op blootstelling aan infectieuze middelen of contact opnemen met bepaalde micro -organismen, hoewel beide systemen meestal samenwerken.
Er wordt gezegd dat het een specifiek systeem is omdat het optreedt als reactie op gedefinieerde determinanten en wordt gemedieerd door extreem gespecialiseerde cellen die ook het vermogen hebben om "te onthouden" en sneller te reageren en met meer "kracht" of "effectiviteit" op herhaalde tentoonstellingen aan dezelfde binnenvallende agent.
Humorale immuniteit is een van de subcategorieën van adaptieve of specifieke immuniteit, die ook wordt geclassificeerd als cellulaire immuniteit. Beide soorten antwoorden verschillen van elkaar volgens het onderdeel van het immuunsysteem dat deelneemt.
[TOC]
Theorie
De theorie van humorale immuniteit, die het product was van intense jaren van onderzoek en debat, stelt voor dat immuniteit wordt gemedieerd door stoffen die aanwezig zijn in lichaamsvloeistoffen of "stemmingen".
Deze theorie is ontwikkeld door veel wetenschappers, die onafhankelijk bestudeerden en beschreven hebben van veel van de effectoren die betrokken waren bij dergelijke mechanismen van de respons.
Paul Ehrlich was misschien een van de meest invloedrijke, omdat hij begin 1900 de diepste studies uitvoerde naar de antigeen-antilichaamcomplementariteit.
Een beetje geschiedenis
De gerenommeerde immunoloog Rudolph Virchow, in 1858, stelde vast dat alle lichaamspathologieën te wijten waren aan de storing van de cellulaire elementen die verantwoordelijk zijn voor bescherming, in plaats van de oplosbare "mismatch van een" stemming ".
Kleine meer dan 25 jaar later, in 1884, bracht Eli Metchnikoff de eerste publicatie van de fagocytische theorie aan het licht, die vandaag de hoofdbases van cel -gemedieerde immuniteitstheorie (celimmuniteit) vormde en ondersteunt.
Kan u van dienst zijn: AndroginiaVeel tegenstanders van Metchnikoff probeerden hun verklaringen te "ontkennen" en het was in 1888 toen George Nuttall, die een reeks experimenten uitvoerde die was ontworpen om metchnikoff te testen, vastgesteld dat het serum van normale dieren een "natuurlijke toxiciteit" had voor bepaalde micro -organismen.
Op deze manier werd het in de wetenschappelijke wereld populair dat cellen vrij van cellen van "gezonde" of "vooral geïmmuniseerde" dieren bacteriën konden doden, op een manier die niet nodig was om hun toevlucht te nemen tot de celtheorie om aangeboren immuniteit te verklaren en verworven.
De eerste die experimenteel het bestaan van een humorale immuunrespons te verifiëren, waren Emil von Behring en Shibasaburo Kitasato in het laatste decennium van 1800. Von Behring en Kitasato toonden aan dat de immuunresponsen die werden afgevuurd door difterie en tetanus te wijten waren aan de aanwezigheid van antilichamen tegen exotoxine.
Begin 1900 realiseerden Karl Landsteiner en andere onderzoekers zich dat andere toxines en stoffen van niet -bacteriële oorsprong humorale immuniteit konden veroorzaken.
De term "antilichaam" werd kort na een algemeenheid bedacht om te verwijzen naar die specifieke stoffen die kunnen functioneren als antitoxinen tegen "antigenen".
Het woord antigeen was de term die werd gebruikt om stoffen te definiëren die de productie van humorale antilichamen veroorzaken.
Effectieve mechanismen
Zowel humorale immuunresponsen als cellulaire immuunresponsen worden gemedieerd door een type cellen bekend als lymfocyten.
De hoofdrolspelers van celimmuniteit zijn T -lymfocyten, terwijl het B -lymfocyten zijn die reageren op de aanwezigheid van vreemde antigenen en transformeren in antilichaamproducerende cellen die kenmerkend zijn voor humorale immuniteit.
Humorale immuniteit is het belangrijkste afweermechanisme tegen extracellulaire micro -organismen en andere toxines, ondertussen draagt celimmuniteit bij aan de eliminatie van intracellulaire pathogenen, die "ontoegankelijk" zijn om te herkennen door antilichaam.
Humorale immuunresponsfasen
Naast de cellulaire cellulaire respons kan de humorale respons worden verdeeld in drie fasen: een van herkenning, een andere van activering en een andere realisatie.
De herkenningsfase bestaat uit de vereniging van antigenen tegen specifieke membraanreceptoren op het celoppervlak van volwassen B -lymfocyten.
Antilichamen functioneren als deze "receptoren" en kunnen eiwitten, polysachariden, lipiden en andere "vreemde" extracellulaire stoffen herkennen.
De activeringsfase begint met de proliferatie van lymfocyten na de herkenning van antigenen en gaat door met differentiatie, hetzij in andere effectorcellen die in staat zijn om antigenen te elimineren, of in geheugencellen die snellere responsen kunnen induceren na een nieuwe blootstelling aan dezelfde antigeen.
Tijdens de effectorfase staan lymfocyten die antigeen -eliminatiefuncties uitoefenen bekend als "effectorcellen", hoewel andere cellen meestal deelnemen, die ook deelnemen aan de aangeboren immuunrespons, en dat phagocytn en het elimineren van vreemde middelen.
Het kan u van dienst zijn: het belang van fossielen: 5 fundamentele redenenLymfocyten en antilichamen
Antilichamen geproduceerd door lymfocyten of B -cellen hebben de fysiologische functie van het neutraliseren en elimineren van het antigeen dat hun vorming induceerde en het humorale immuunsysteem kan reageren op veelheid van verschillende antigenen.
B -lymfocyten zijn afkomstig van het beenmerg in reactie op een gedefinieerd antigeen (ze zijn specifiek) en dit gebeurt vóór antigene stimulatie. De specifieke antilichaamuitdrukking veroorzaakt de reacties van proliferatie en differentiatie van meer B B -cellen van antilichaamsecretoren.
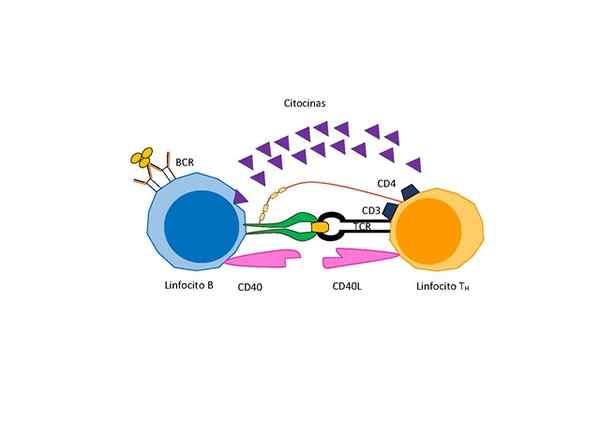
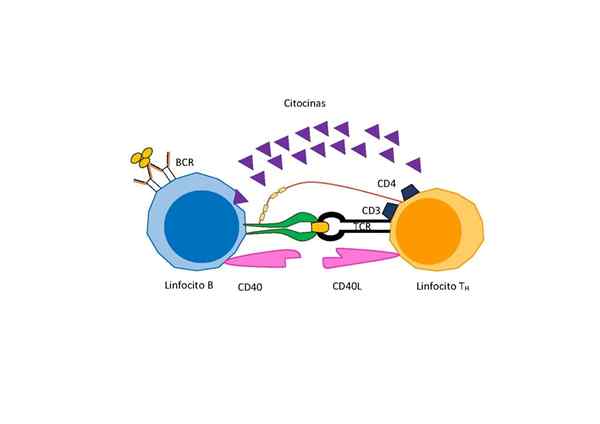 Signalering tussen T -cellen en B -cellen voor de activering van deze laatste (Bron: Manuel Mellina Vicente, via Wikimedia Commons)
Signalering tussen T -cellen en B -cellen voor de activering van deze laatste (Bron: Manuel Mellina Vicente, via Wikimedia Commons) Afhankelijk van de aard van het antigeen is echter een extra signaal nodig voor de differentiatie en proliferatie gegeven door een speciaal type T -lymfocyten genaamd "samenwerkende T -lymfocyten" die geheime activeringsfactoren voor B -cellen geheime.
Immunoglobulinen
Gezien het feit dat ze voornamelijk in bloedvloeistoffen zijn, worden antilichamen geproduceerd door B -cellen immunoglobulines genoemd. Deze eiwit natuurmoleculen hebben twee zware glycoproteïne ketens en twee lichtgewicht aan elkaar verbonden door disulfurbruggen (S-S).
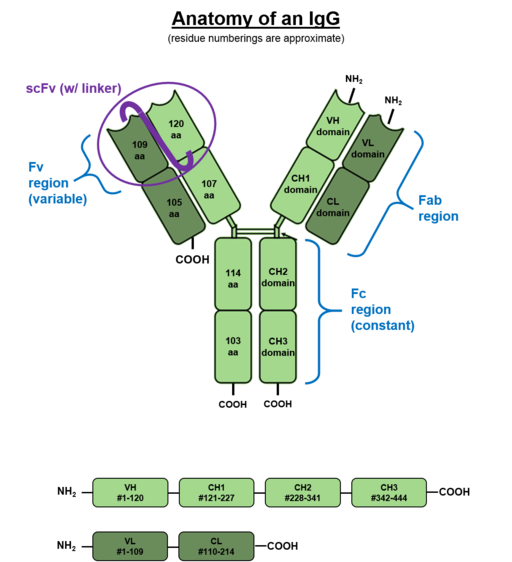 Structuur van een immunoglobuline G (IgG) (Bron: W: Gebruiker: Ajvincelli [Public Domain] via Wikimedia Commons)
Structuur van een immunoglobuline G (IgG) (Bron: W: Gebruiker: Ajvincelli [Public Domain] via Wikimedia Commons) Lichte ketens staan bekend als "kappa" en "lambda", maar er zijn 5 soorten zware ketens die gamma (g), mu (m), alpha (a), delta (d) en epsilon (e) worden genoemd.
De combinatie van lichte en zware ketens eindigt met de vorming van IgG, IgM, IgD en IgE -immunoglobulines. Het meest voorkomende antilichaam in zoogdierserum in IgG -immunoglobuline (ongeveer 70%).
Elke keten van een antilichaam heeft een amino -terminaaluiteinde en een ander terminaalcarboxyl. Het gedeelte dat in staat is om de antigenen aan te sluiten, bevindt zich aan het amino -terminale uiteinde, maar het terminale carboxylgebied is degene die biologische activiteit bepaalt.
Humorale immuunrespons
Het terminale carboxylgebied van antilichamen van het IgG -type wordt specifiek herkend door fagocytische cellen zoals neutrofielen en macrofagen, die hiervoor speciale receptoren hebben.
Deze herkenning impliceert contact tussen de ontvanger en het antilichaam en het is deze unie die fagocytose en afbraak van antigenen in fagocytische cellen vergemakkelijkt.
In tegenstelling tot IgG worden de andere soorten immunoglobulines niet gevonden in secreties en weefsels. Dit zijn echter even gereedschap bij het uitlokken van de immuunrespons.
IgM -immunoglobulinen (10% van de serum -immunoglobulines) zijn krachtige complementsysteemactivatoren, dus ze werken in de lysis van antigenen en verhoogde resistentie.
Kan u van dienst zijn: 12 vragen over de opwarming van de aardeIgA -immunoglobulines (20% van de serum -immunoglobulines) worden geproduceerd in lymfoïde weefsels en worden verwerkt en getransporteerd naar longslijmvliezen en maagdarmkanaal. Deze werken bij de neutralisatie van virussen en andere antigenen die de slijmvlies binnenkomen.
De IGD is gekoppeld aan B -lymfocyten en functioneert als een antigenen -ontvanger, terwijl het IgE (bekend als het allergische antilichaam) is gekoppeld aan het oppervlak van mestcellen en basofielen via specifieke receptoren. Beide immunoglobulinen zijn in een zeer lage concentratie in serum.
Voorbeelden
De antilichamen geproduceerd door de belangrijkste effectoren van de humorale immuunrespons (lymfocyten B) hebben het vermogen om verschillende responsmechanismen te "induceren" of "activeren" tegen verschillende soorten bedreigingen.
IgG -immunoglobulinen zijn bijvoorbeeld activatoren van wat bekend staat als de "complement waterval", die werkt aan de neutralisatie van virale deeltjes, waardoor hun vereniging wordt voorkomen om cellen te hosten.
Tijdens de zwangerschap draagt de moeder antilichamen over naar de foetus door de trofoblastische cellen van de placenta, die hoge affiniteitsreceptoren hebben voor het einde van immunoglobulines terminale carboxyl zoals IgG zoals IgG zoals IgG zoals IgG.
De humorale reactie op bacteriën met "capsules" bestaande uit polysachariden wordt gemedieerd door immunoglobuline M, die fagocytose van deze micro -organismen bevordert.
Een ander belangrijk voorbeeld van humorale immuniteit is de systemische reactie op parasieten, waarbij het IgE hun vernietiging door eosinofiele cellen "richt".
Referenties
- Abbas, een., Lichtman, een., & Armen, j. (1999). Cellulaire en moleculaire immunologie (3e ed.)). Madrid: McGraw-Hill.
- Carroll, m. C., & Isenman, D. EN. (2012). Regulering van humorale immuniteit door complement. Immuniteit, 37(2), 199-207.
- Kindt, T., Goldsby, r., & Osborne, B. (2007). Kuby -immunologie (6e ed.)). Mexico D.F.: McGraw-Hill Inter-American uit Spanje.
- Klein, t. (2007). De verworven immuunrespons. In Xpharm: de uitgebreide farmacologie -referentie (PP. 1-5).
- Lishner, h., & Digeorge, a. (1969). Rol van de thymus in humoristische immuniteit. Het Lancet, 2, 1044-1049.
- Medzhitov, R., & Janeway, c. (2000). Aangeboren immuniteit. The New England Journal of Medicine, 338-344.
- Merlo, l. M. F., & Mandik-Nayak, L. (2013). Adaptieve Immunch: B -cellen en antilichamen. In Immunotherapie kanker: immuunonderdrukking en tumorgroei: tweede editie (PP. 25-40).
- Silverstein, a. M. (1979). Geschiedenis van immunologie. Cellulaire versus humor -immuniteit: determinanten en verwakeling van een epische 19e -eeuwse strijd. Cellulaire immunologie, 48(1), 208-221.
- Steinman, r. M. (2008). Aangeboren aan adaptief immuun verbinden door dendritische cellen. In Aangeboren immuniteit voor longinfectie (PP. 101-113).
- Tan, t. T., & Cousens, l. M. (2007). Humorale immuniteit, ontsteking en kanker. Huidige mening in immunologie, 19(2), 209-216.
- Twigg, h. L. (2005). Humorale immuunafweer (antilichamen): recente vorderingen. Proceedings of the American Thoracic Society, 2(5), 417-421.
- Wherry, E. J., & Masopust, D. (2016). Adaptieve immuniteit: neutralisatie, elimineren en onthouden voor de volgende keer. In Virale pathogenese: van basics tot systeembiologie: derde editie (PP. 57-69).