

Oxidatieve fosforyleringsfasen, functies en remmers
- 1376
- 118
- Hugo Crooks
De oxidatieve fosforylering Het is een proces waarbij ATP -moleculen worden gesynthetiseerd uit ADP en PJe (Anorganisch fosfaat). Dit mechanisme wordt uitgevoerd door bacteriën en eukaryotische cellen. In eukaryotische cellen wordt fosforylering uitgevoerd in de mitochondriale matrix van niet -fotosynthetische cellen.
ATP -productie wordt geleid door de overdracht van elektronen uit de NADH- of FADH -co -enzymen2 te2. Dit proces vertegenwoordigt de grootste energieproductie in de cel en is afgeleid van de afbraak van koolhydraten en vetten.
Bron: Robot8a [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] De energie die is opgeslagen in de belasting- en pH -gradiënten, ook bekend als protonische motoriekracht, maakt het mogelijk dat dit proces wordt uitgevoerd. De gegenereerde protongradiënt, zorgt ervoor dat het externe deel van het membraan een positieve belasting heeft vanwege de concentratie van protonen (h+) en de mitochondriale matrix is negatief.
[TOC]
Waar oxidatieve fosforylering optreedt?
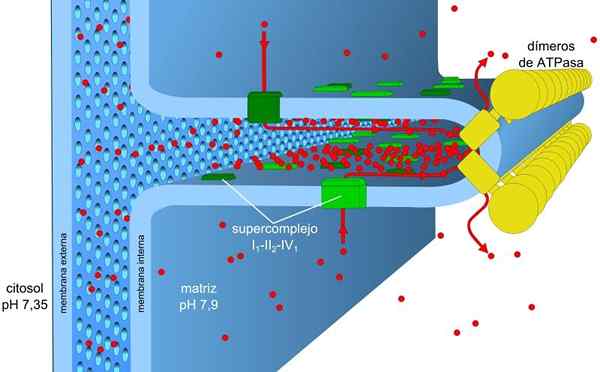
Elektronentransport en oxidatieve fosforyleringsprocessen zijn geassocieerd met een membraan. In prokaryoten worden deze mechanismen uitgevoerd door het plasmamembraan. In eukaryotische cellen worden ze geassocieerd met het mitochondria -membraan.
Het aantal mitochondriën dat in de cellen wordt gevonden, varieert volgens het celtype. Bij bijvoorbeeld zoogdieren missen erytrocyten deze organellen, terwijl andere celtypen, zoals spiercellen, er miljoenen van kunnen hebben.
Het mitochondriale membraan bestaat uit een eenvoudig extern membraan, een enigszins complexer intern membraan, en in het midden daarvan de intermembranale ruimte, waar veel enzymen afhankelijk zijn van ATP bevinden.
Het buitenmembraan bevat een eiwit genaamd porina dat de kanalen vormt voor de eenvoudige diffusie van kleine moleculen. Dit membraan is verantwoordelijk voor het handhaven van de structuur en vorm van de mitochondria.
Het interne membraan heeft een grotere dichtheid en is rijk aan eiwitten. Het is ook waterdicht voor moleculen en ionen, dus om het over te steken, hebben ze intermembranale eiwitten nodig die ze transporteren.
Binnen de matrix strekken vouwen van het interne membraan zich uit, waardoor ruggen vormen waarmee het een groot gebied in een klein volume kan hebben.
Cell Energy Central
De mitochondria wordt beschouwd als het Cellular Energy Center. Daarin zijn de enzymen die betrokken zijn bij de processen van de cytrinezuurcyclus, oxidatie van vetzuren en redox -enzymen en eiwitten van elektronentransport en PhD -fosforylering.
De concentratiegradiënt van de protonen (pH -gradiënt) en de gradiënt van belastingen of elektrische potentiaal in het interne membraan van de mitochondriën, zijn de oorzaak van de Prottonische motoriekracht. De kleine permeabiliteit van het interne membraan voor ionen (anders dan h+) stelt de mitochondria in staat om een stabiele spanningsgradiënt te hebben.
Elektronisch transport, protonen pompen en ATP die tegelijkertijd in de mitochondria verkrijgen, dankzij de Protonic Motor Force. De pH -gradiënt handhaaft zure omstandigheden in intermembraan en mitochondriale matrix met alkalische omstandigheden.
Voor elke twee elektronen overgebracht naar de O2 Ongeveer 10 protonen worden gepompt door het membraan, waardoor een elektrochemische gradiënt ontstaat. De energie die in dit proces is losgemaakt, wordt geleidelijk geproduceerd door elektronen door de transportketen te passeren.
Fasen
De energie die vrijkomt tijdens oxide-reductiereacties van NADH en FADH2 Het is aanzienlijk hoog (ongeveer 53 kcal/mol voor elk paar elektronen), om te worden gebruikt bij de productie van ATP -moleculen, moet het geleidelijk worden geproduceerd met de doorgang van elektronen door transporters.
Deze zijn georganiseerd in vier complexen in het interne mitochondriale membraan. De koppeling van deze reacties op ATP -synthese wordt uitgevoerd in een vijfde complex.
Kan u van dienst zijn: B -lymfocyten: kenmerken, structuur, functies, typenElektronentransportketen
NADH draagt een paar elektronen over die de complex I van de elektrontransportketen binnenkomen. De elektronen worden overgebracht naar het mononucleotide van flavina, en vervolgens naar de ubiquinon (co-enzym q) door een transportbeurt. Dit proces geeft een grote hoeveelheid energie vrij (16,6 kcal/mol).
Ubiquinona transporteert elektronen door het membraan naar complex III. In dit complex gaan de elektronen door cytochromen B en C1 Dankzij een transporter van ijzer-suiker.
Van complex III passeren elektronen naar het IV -complex (cytochroom C -oxidase), overgebracht door cytochroom C (perifeer membraaneiwit). In het IV -complex gaan de elektronen door een paar koperionen (Cunaar2+), dan naar Cytochrome Cnaar, dan naar een ander paar koperionen (CuB2+) en van dit tot cytochroom tot3.
Ten slotte worden elektronen overgebracht naar O2 die de laatste acceptor is en een watermolecuul vormt (h2O) Voor elk ontvangen paar elektronen. De doorgang van elektronen van complex IV naar O2 Het genereert ook veel vrije energie (25,8 kcal/mol).
Succinato coq reductase
Het II -complex (coq -reductase succinate) ontvangt een paar elektronen uit de citroenzuurcyclus, vanwege de oxidatie van een succinaatmolecuul tot fumaraat. Deze elektronen worden overgebracht naar de rage en gaan vervolgens door een ijzerzufre-groep, naar de Ubiquinona. Vanuit dit co -enzyme gaan ze naar Complex III en volgen de eerder beschreven route.
De energie die wordt vrijgegeven in de elektronenoverdrachtsreactie op de rage is niet voldoende om de protonen door het membraan te stimuleren, zodat er in deze stap van de ketting geen Prottonische motoriekracht is, en bijgevolg geeft de FADH minder h+ dan de NADH.
Energiekoppeling of transductie
De energie die is gegenereerd in het eerder beschreven elektronentransportproces, moet worden gebruikt voor ATP -productie, reactie gekatalyseerd door het ATP -synthase of complex enzym V. Het behoud van dergelijke energie staat bekend als energiekoppeling en het mechanisme is moeilijk te karakteriseren.
Verschillende hypothesen zijn beschreven om deze energietransductie te beschrijven. De best geaccepteerde is de chemosmotische koppelingshypothese, hieronder beschreven.
Chemosmotische koppeling
Dit mechanisme stelt voor dat de energie die wordt gebruikt voor ATP -synthese afkomstig is van een protonische gradiënt in celmembranen. Dit proces komt tussenbeide in mitochondriën, chloroplasten en bacteriën en is gekoppeld aan elektronentransport.
De I en IV -complexen van elektronisch transport werken als protonpompen. Deze lijden aan conformationele veranderingen waarmee ze de protonen naar de intermembranale ruimte kunnen pompen. In het IV -complex voor elk paar elektronen worden twee protonen buiten het membraan gepompt en er blijven er nog twee in de matrixvorming h2OF.
Ubiquinona in complex III accepteert protonen van complexen I en II en bevrijdt ze buiten het membraan. I- en III -complexen staan elk de doorgang van vier protonen toe voor elk paar getransporteerde elektronen.
De mitochondriale matrix heeft een lage concentratie van protonen en negatieve elektrische potentiaal, terwijl de intermembranale ruimte de omgekeerde omstandigheden presenteert. De stroom van protonen door dit membraan impliceert de elektrochemische gradiënt die de noodzakelijke energie (± 5 kcal/mol door proton) opslaat voor ATP -synthese.
ATP -synthese
Het ATP -synthetase -enzym is het vijfde complex dat betrokken is bij oxidatieve fosforylering. Is verantwoordelijk voor het profiteren van de energie van de elektrochemische gradiënt om ATP te vormen.
Dit transmembraal eiwit bestaat uit twee componenten: f0 en f1. De component f0 maakt de teruggave van de protonen naar de mitochondriale matrix mogelijk als een kanaal en de F1 katalyseert de synthese van de ATP via ADP en PJe, De energie van genoemde rendement gebruiken.
Kan u van dienst zijn: Membraantransporteur Eiwitten: functies en typenHet ATP -syntheseproces vereist een structurele verandering in F1 en de assemblage van componenten f0 en f1. De translocatie van protonen via F0 Het veroorzaakt conformationele veranderingen in drie subeenheden van F1, waardoor dit kan fungeren als een rotatie -engine, die de vorming van de ATP regisseert.
De subeenheid die verantwoordelijk is voor de Unie van ADP met PJe gaat van een zwakke toestand (l) naar een actieve (t). Wanneer de ATP wordt gevormd, gaat een tweede subeenheid door naar een open toestand (O) die de afgifte van deze molecuul mogelijk maakt. Nadat de ATP is vrijgegeven, gaat deze subeenheid van de open staat over naar een inactieve staat (L).
De ADP- en P -moleculenJe Ze sluiten zich aan bij een subeenheid die is gegaan van een staat of de staat L.
Producten
De elektronentransportieketen en fosforylering produceren ATP -moleculen. NADH -oxidatie produceert ongeveer 52,12 kcal/mol (218 kJ/mol) vrije energie.
De globale reactie voor NADH -oxidatie is:
NADH+1⁄2 of2 +H+ ↔ H2O+NAD+
De overdracht van elektronen van NADH en FADH2 Het komt voor door verschillende complexen, waardoor de verandering van vrije energie Ag ° kan worden verdeeld in kleinere "pakketten", die zijn gekoppeld aan ATP -synthese.
De oxidatie van een NADH -molecuul genereert de synthese van drie ATP -moleculen. Terwijl de oxidatie van een FADH -molecuul2 Het is bevestigd aan de synthese van twee ATP.
Deze co -enzymen komen uit de processen van glycolyse en citroenzuurcyclus. Voor elk afgebroken glucosemolecuul eindigen 36 of 38 ATP -moleculen, afhankelijk van de locatie van de cellen. In de hersenen en skeletspier worden 36 ATP geproduceerd terwijl 38 ATP wordt geproduceerd in spierweefsel.
Functie
Alle organismen, eencellig en meercellig, hebben een minimale energie in hun cellen nodig om de processen erin uit te voeren, en behouden op hun beurt vitale functies in het volledige lichaam.
Metabole processen vereisen dat energie wordt uitgevoerd. De meeste bruikbare energie wordt verkregen door de afbraak van koolhydraten en vetten. Deze energie is afgeleid van het oxidatieve fosforyleringsproces.
Oxidatieve fosforyleringscontrole
De ATP -gebruikssnelheid in de cellen, regelt de synthese van hetzelfde en op zijn beurt, vanwege de koppeling van oxidatieve fosforylering met de elektronentransportketen, reguleert het in het algemeen ook het elektronische transportsnelheid.
Oxidatieve fosforylering heeft een strikte controle die ervoor zorgt dat ATP niet sneller wordt gegenereerd dan het wordt geconsumeerd. Er zijn bepaalde stappen in het elektrontransportproces en de gekoppelde fosforylering die de energieproductiesnelheid reguleren.
Gecoördineerde controle van ATP -productie
De belangrijkste energieproductieroutes (cel ATP) zijn glycolyse, de cyclus van citroenzuur en oxidatieve fosforylering. Gecoördineerde controle van deze drie processen reguleert de ATP -synthese.
Fosforyleringscontrole door de ATP -massa -werkingsverhouding hangt af van de precieze bijdrage van elektronen in de transportketen. Dit hangt op zijn beurt af van de relatie [NADH]/[NAD+] dat wordt bewaard verhoogd door de werking van glycolyse en de citroenzuurcyclus.
This coordinated control is carried out by regulating glycolysis control points (Citrate inhibited PFK) and the cytric acid cycle (dehydrogenase pyruvate, cutease citrate, itchitrate dehydrogenase and α-zetoglutarate dehydrogenase).
Controle door acceptor
Het IV -complex (cytochroom C -oxidase) is een enzym gereguleerd door een van zijn substraten, dat wil zeggen dat de activiteit wordt geregeld door de verminderde cytochroom C (C (C2+), die op zijn beurt in evenwicht is met de relatie tussen de concentraties tussen [NADH]/[NAD+] en de massa -actieverhouding van [ATP]/[ADP] + [PJe].
Het kan u van dienst zijn: Tone: Kenmerken en functiesHoe hoger de [NADH]/[NAD+en verlagen de [ATP]/[ADP]+[PJe], meer concentratie zal cytochroom zijn [c2+] en groter zal de activiteit zijn van complex IV. Dit wordt bijvoorbeeld geïnterpreteerd als we organismen vergelijken met verschillende rustactiviteiten en hoge activiteiten.
Bij een individu met hoge fysieke activiteit, de consumptie van ATP en dus de hydrolyse ervan tot ADP + PJe Het zal erg hoog zijn, waardoor een verschil wordt gegenereerd in de massa -actieverhouding die een toename van [C veroorzaakt2+en daarom een toename van de ATP -synthese. In een rustend persoon treedt de omgekeerde situatie voor.
Uiteindelijk neemt de snelheid van oxidatieve fosforylering toe met de concentratie van ADP binnen de mitochondria. Deze concentratie hangt af van ADP-ATP-translocators die worden beschuldigd van het transport van adenine-nucleotiden en PJe Van cytosol tot de mitochondriale matrix.
Decacoplants agenten
Oxidatieve fosforylering wordt beïnvloed door bepaalde chemische middelen, waardoor elektronisch transport kan doorgaan zonder de fosforylering van de ADP, waardoor de productie en het behoud van energie ontkoppelt.
Deze middelen stimuleren de mitochondria -zuurstofverbruiksnelheid in afwezigheid van ADP, wat ook een toename van ATP -hydrolyse veroorzaakt. Ze werken door een tussenpersoon te elimineren of een staat van energie te breken uit de elektronentransportieketen.
Het 2.4-dinitrofenol, een zwak zuur dat door de mitochondriale membranen gaat, is verantwoordelijk voor het dissiperen van de protonengradiënt, omdat ze zich aan de zure kant bij elkaar geven en aan de basiszijde vrijgeven.
Deze verbinding werd gebruikt als een "afslankpil" omdat het bleek dat het een toename van de ademhaling veroorzaakte, daarom een toename van de metabole snelheid en bijbehorende gewichtsverlies. Er werd echter aangetoond dat het negatieve effect ervan zelfs de dood zou kunnen veroorzaken.
De dissipatie van de protonische gradiënt veroorzaakt warmte. Bruine vetweefselcellen gebruiken ontkoppeling, hormonaal gecontroleerd, om warmte te produceren. Hibernate zoogdieren en pasgeborenen die haar missen, bestaan uit dit weefsel dat hen dient als een soort thermische deken.
Remmers
Remmende verbindingen of middelen voorkomen zowel de consumptie van of2 (elektronisch transport) zoals bijbehorende oxidatieve fosforylering. Deze agenten vermijden de vorming van ATP door het gebruik van energie geproduceerd in elektronisch transport. Daarom stopt de transportketen omdat een dergelijk energieverbruik niet is gerangschikt.
Oligomicine -antibioticum werkt als een fosforylatieremmer bij veel bacteriën, waardoor ADP -stimulatie van ATP -synthese wordt voorkomen.
Er zijn ook ionoforeuze middelen, die vetoplosbare complexen vormen met kationen zoals k+ en NA+, en passeer het mitochondriale membraan met deze kationen. De mitochondria gebruikt vervolgens energie geproduceerd in elektronisch transport om kationen te pompen in plaats van ATP te synthetiseren.
Referenties
- Alberts, B., Bray, D., Hopkin, K., Johnson, a., Lewis, J., Raff, m., Roberts, K. & Walter, p. (2004). Essentiële celbiologie. New York: Garland Science.
- Cooper, G. M., Hausman, r. EN. & Wright, n. (2010). De cel. (PP. 397-402). Marbán.
- Devlin, T. M. (1992). Textbook of Biochemistry: met klinische correlaties. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, c. M. (2008). Biochemie. Thomson Brooks/Cole.
- Lodish, h., Darnell, J. EN., Berk, een., Kaiser, c. NAAR., Krieger, m., Scott, m. P., & Matsudaira, p. (2008). Molleculaire celbiologie. Macmillan.
- Nelson, D. L., & Cox, m. M. (2006). Lehninger -principes van biochemie 4e editie. Ed Omega. Barcelona.
- VOET, D., & Voet, J. G. (2006). Biochemie. ED. Pan -American Medical.
- « Euglenophyta -kenmerken, reproductie, voeding, classificatie
- Glucosides training, functie en typen/groepen »