

Flagelina -structuur en functies
- 1979
- 265
- Ernesto McKenzie
De Vlag Het is een eiwit van de gloeidraad, een structuur die deel uitmaakt van de plaag van bacteriën. De overgrote meerderheid van de bacteriën heeft slechts één type plaag. Sommigen hebben echter meer dan twee.
De moleculaire grootte van dit eiwit varieert tussen 30 kDa en 60 kDa. In Enterobacteria is de moleculaire grootte bijvoorbeeld groot, terwijl in bepaalde sweetacuícolas bacteriën klein is.
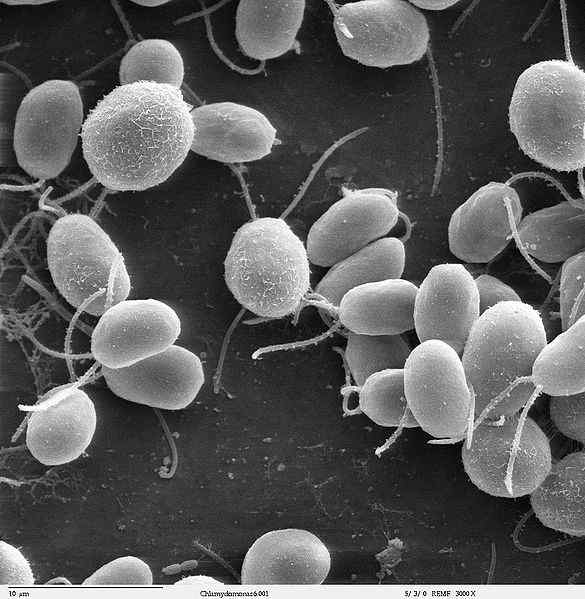
Bron: Dartmouth Electron Microscope Facility, Dartmouth College [Public Domain] Flagelina is een virulentiefactor die hechting en invasie aan gastheercellen mogelijk maakt. Bovendien is het een krachtige activator van vele soorten cellen die betrokken zijn bij de aangeboren en adaptieve immuunrespons.
[TOC]
Flagelo en mobiliteit Ultrastructuur
De gesel is verankerd naar het celoppervlak. Het bestaat uit drie delen: 1) de gloeidraad, die zich uitstrekt van het oppervlak van de cel en een holle en rigide cilindrische structuur is; 2) het basale lichaam, dat is ingebed in de wanden van wand- en celmembraan, die verschillende ringen vormt; en 3) de haak, een gebogen korte structuur die het basale lichaam bij de gloeidraad verbindt.
Het basale lichaam is het meest complexe deel van de plaag. In gram negatieve bacteriën heeft het vier ringen verbonden met een centrale kolom. In de positieve gram heeft het twee ringen. De rotatiebeweging van de gesel vindt plaats in het basale lichaam.
De locatie van de flagella op het oppervlak van de bacterie varieert sterk tussen organismen, in staat zijn: 1) monric, met slechts één plaag; 2) polair, met twee of meer; of 3) peritico, met veel zij flagella. Er zijn ook endoflagelo's, zoals in de spirocheten, die zich in de PLAPSMIC -ruimte bevinden.
Helicobacter Pylori Het is erg mobiel omdat het zes tot acht unipolaire gejaag heeft. Een pH -gradiënt door slijm laat het toe H. pylori Zich oriënteren en zich vestigen in een gebied grenzend aan epitheelcellen. Pseudomonas Het heeft een polaire gesel, die chemiotaxis voor suikers vertoont en wordt geassocieerd met virulentie.
Fuseline structuur
Een kenmerk van de chattering-eiwitsequentie is dat de N-terminale en C-terminale regio's sterk bewaard zijn, terwijl het centrale gebied zeer variabel is tussen soorten en ondersoorten van hetzelfde geslacht. Deze hypervariabiliteit is verantwoordelijk voor honderden serotypes van Salmonella spil.
Kan u van dienst zijn: Serratia MarcescensFlagelina -moleculen interageren met elkaar via de terminale gebieden en polymeriseren het vormen van een gloeidraad. Hierin bevinden de terminale gebieden zich in het interieur van de cilindrische structuur van de gloeidraad, terwijl de centrale wordt blootgesteld.
In tegenstelling tot tubulinefilamenten die zijn gedepolimereerd in afwezigheid van zouten, zijn bacteriën zeer stabiel in water. Ongeveer 20.000 tubuline -subeenheden vormen een gloeidraad.
In de gloeidraad van H. pylori En Pseudomonas aeruginosa Twee soorten gesel en flab worden gepolymeriseerd, gecodeerd door het FLIC -gen. FLAA is heterogeen en zijn onderverdeeld in verschillende subgroepen, met moleculaire massa's die variëren tussen 45 en 52 kDa. Flab is homogeen met een moleculaire massa van 53 kDa.
Vaak worden flageline -lysineresten gemethyleerd. Bovendien zijn er andere modificaties zoals FLAA -glysilatie en fosforylering van flab -tyrosineverval, waarvan de functies respectievelijk virulentie en exportsignaal zijn.
Flagerie filamentgroei in bacteriën
De plaag van bacteriën kan experimenteel worden ervaren, waardoor de regeneratie de regeneratie kan bestuderen. Flagelina -subeenheden worden getransporteerd door het interne gebied van deze structuur. Wanneer ze het einde bereiken, voegen de subeenheden spontaan toe met behulp van een eiwit ("cap -eiwit") genaamd hap2 of floid.
De synthese van de gloeidraad vindt plaats door zijn eigen vergadering; dat wil zeggen, de polymerisatie van de plaag vereist geen enzymen of factoren.
De informatie voor de vergadering van de gloeidraad is te vinden in de subeenheid zelf. Aldus polymeriseren flagelina -subeenheden het vormen van elf protofilamenten, die een complete vormen.
Flagelina -synthese van P. aeruginosa En Proteus Mirabilis Het wordt geremd door antibiotica zoals erytromycine, clarithromycine en azithromycine.
Flagelina als een immuunsysteemactivator
De eerste studies toonden aan dat de gesel, bij subnomolaire concentraties, van Salmonella, Het is een krachtige cytokine -inductor in een promocitale cellijn.
Kan u van dienst zijn: zoosporenVervolgens werd aangetoond dat de inductie van de pro -inflammatoire respons een interactie impliceert tussen de plaag en de oppervlakteceptoren van het aangeboren immuunsysteemcellen.
De oppervlakteceptoren die interageren met flageline zijn die van het tol-5 (TLR5) type. Vervolgens toonden studies met recombinant gesel aan dat, wanneer het het hypervariabele gebied ontbrak, het geen immuunrespons kon induceren.
TLR5 zijn aanwezig in immuunsysteemcellen, zoals lymfocyten, neutrofielen, monocyten, macrofagen, dendritische cellen, epitheelcellen en lymfatische knobbeltjes. In de darm reguleert TLR5 de samenstelling van de microbiota.
Gram negatieve bacteriën gebruiken meestal het type-III-secretoire systeem om de plaag naar het cytoplasma van de gastheercel te translocaliseren, dat een reeks intracellulaire gebeurtenissen veroorzaakt. Aldus wordt de flageline in de intracellulaire omgeving erkend door eiwitten van de NAIP -familie (een remmer van apoptose/nlr -familie).
Vervolgens interageert het Flagelina-NAIP5/6-complex met de ontvanger van het NOW-type, dat de reactie van de host op infectie en schade genereert.
Vlagelina en planten
Planten herkennen dit eiwit via de weg Detectie 2 van de gesel (fls2). De laatste is een kinasereceptor in leucinerepetities en is een tegenhanger van TLR5. FLS ”interageert met de N-terminale regio van LA Flagelina.
De vereniging van flageline tot FLS2 produceert fosforylering van de MAP -kinase -weg, die culmineert met de synthese van eiwitten die bescherming bemiddelen tegen schimmelinfectie en bacteriën.
In sommige Solanáceas -planten kan de gesel ook lid worden van de FLS3 -receptor. Op deze manier worden deze beschermd tegen pathogenen die de verdediging ontwijken gemedieerd door FLS2.
Vlagelina als adjuvans
Een adjuvans is een materiaal dat de cel of humorale respons op een antigeen verhoogt. Omdat veel vaccins een slechte immuunrespons produceren, is het noodzakelijk om goede adjuvantia te hebben.
Kan u dienen: heterotrofe bacteriën: kenmerken en voorbeelden van soortenTalrijke studies toonden de effectiviteit van gesel als adjuvans. Deze onderzoeken bestonden uit het gebruik van recombinant flageline in vaccins, geëvalueerd door diermodellen. Dit eiwit wordt echter nog steeds overwonnen door fase I van klinische onderzoeken.
Onder de bestudeerde recombinante flageline's zijn: Flagelina-Epitope 1 van hematoglutinine van het griepvirus; Flagelina-epitoop Schistosoma mansoni; Flageline-stabiele warmte om te verwarmen EN. coli; Flagelina -proteína 1 van het oppervlak van Plasmodium; en vlagelina-eiwit van het Nile-virusverpakking, onder andere recombinanten.
Er zijn enkele voordelen om flageline te gebruiken als adjuvans in menselijke gebruiksvaccins. Deze voordelen zijn als volgt:
1) Het is effectief bij zeer lage doses.
2) Ze stimuleren het IgE -antwoord niet.
3) U kunt de volgorde van een ander adjuvans, Ag, in de plaagsequentie invoegen zonder het flagelijnsignaal te beïnvloeden via TLR5.
Ander gebruik van flageline
Omdat de flagelinegenen een grote variatie vertonen, kunnen worden gebruikt om specifieke detecties uit te voeren of de identificatie van soorten of stammen te bereiken.
De combinatie van PCR/RFLP is bijvoorbeeld gebruikt om de verdeling en het polymorfisme van flageline -genen in geïsoleerd te bestuderen EN. coli Uit Noord -Amerika.
Referenties
- Hajam, ik. NAAR., Dar, p. NAAR., Shahnawaz, ik., Jaume, J. C., Lees, J. H. 2017. Bacteriële flallin - een vermogensimmunomodulerend middel. Experimentele en moleculaire geneeskunde, 49, E373.
- Kawamura-Sato, K., Inuma, en., Hasegawa, T., Horii, t., Yamashino, T., Ohta, m. 20000000000000000000. Effect van subinhibitorry -concentraties van macroliden op expressie van flamelline in Pseudomonas aeruginosa En Proteus Mirabilis. Antimicrobiële middelen en chemotherapie, 44: 2869-2872.
- Mizel, s. B., Bates, J. T. 2010. Flallin als een attjuvant: cellulaire mechanismen en potentieel. Journal of Immunology, 185, 5677-5682.
- Prescott, l. M., Harley, J. P., Klain, s. D. 2002. Microbiologie. Mc Graw-Hill, New York.
- Schaechter, m. 2009. Het bureau enyclopedie van microbiologie. Academic Press, San Diego.
- Winstanley, c., Morgan, a. W. 1997. Het bacteriële flamelline -gen als biomarker voor detectie, populatiegenetica en epidemiologische analyse. Microbiologie, 143, 3071-3084.