

Typen van celdeling, processen en belang
- 2951
- 230
- Irving McClure I
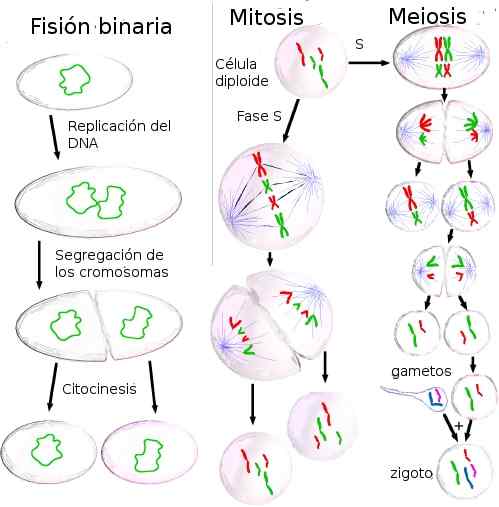
De cellulaire divisie Het is het proces dat alle levende organismen laat groeien en zich voortplanten. In prokaryoten en eukaryoten zijn het resultaat van celdeling dochtercellen die dezelfde genetische informatie hebben als de oorspronkelijke cel. Dit gebeurt omdat, voorafgaand aan de divisie, de informatie in het DNA wordt verdubbeld.
In prokaryoten gebeurt divisie door binaire splijting. Het genoom van de meeste prokaryoten is een cirkelvormig DNA -molecuul. Hoewel deze organismen geen kern hebben, is het DNA in een verdichte vorm genaamd nucleoid, die verschilt van het cytoplasma dat eromheen omringt.
Bron: Retama [CC BY-SA 3.0 (http: // creativeCommons.Org/licenties/by-sa/3.0/]] In eukaryoten gebeurt verdeling door mitose en meiose. Het eukaryotische genoom bestaat uit grote hoeveelheden georganiseerd DNA in de kern. Deze organisatie is gebaseerd op DNA -verpakkingen met eiwitten, die chromosomen vormen, die honderden of duizenden genen bevatten.
De zeer diverse eukaryoten, zowel eencellige als metazoaries, hebben levenscycli die mitose en meiose afwisselen. Deze cycli zijn die met: a) gammatische meiose (dieren, sommige schimmels en algen), b) cygotische meiose (sommige schimmels en protozoa); en c) Afwisseling tussen gamatische en cygotische meiose (planten).
[TOC]
Jongens
Celdeling kan zijn door binaire splijting, mitose of meiose. Het volgende beschrijft elk van de processen die betrokken zijn bij dit soort celdeling.
Binaire splijting
 Procariotische splijting, binaire splijting, is een aseksuele vorm van reproductie.
Procariotische splijting, binaire splijting, is een aseksuele vorm van reproductie. Binaire splijting bestaat uit de verdeling van de cel die aanleiding geeft tot twee dochtercellen, elk met een identieke kopie van het oorspronkelijke cel -DNA.
Vóór de verdeling van de prokaryotische cel vindt de replicatie van DNA plaats, die begint op een specifieke plaats van het DNA met dubbele keten, de oorsprong van de replicatie genoemd. Replicatie -enzymen bewegen naar beide richtingen van de oorsprong en produceren een kopie van elk van de DNA -ketens met dubbele keten.
Na DNA -replicatie verlengt de cel en wordt het DNA in de cel gescheiden. Onmiddellijk begint een nieuw plasmamembraan in het midden van de cel te groeien, waardoor een septum wordt gevormd.
Dit proces wordt vergemakkelijkt door het FTSZ -eiwit, dat evolutionair zeer bewaard is in prokaryoten, waaronder de Archaea. Ten slotte is de cel verdeeld.
Celcyclus en mitose
De fasen waardoor een eukaryotische cel uit twee opeenvolgende celdelingen bekend staat als een celcyclus. De duur van de celcyclus varieert van enkele minuten tot maanden, afhankelijk van het celtype.
De celcyclus is verdeeld in twee fasen, namelijk fase M en interface. Twee processen, mitose genoemd en cytokinese treden op in de M -fase. Mitose bestaat uit nucleaire divisie. Hetzelfde aantal en soorten chromosomen die aanwezig zijn in de oorspronkelijke kern zijn te vinden in de kinderen kernen. Somatische cellen van meercellige organismen worden gedeeld door mitose.
Cytokinese bestaat uit de cytoplasma -deling om dochtercellen te vormen.
De interface heeft drie fasen: 1) G1, de cellen groeien en brengen het grootste deel van hun tijd door in deze fase; 2) s, genoomduplicatie; en 3) G2, replicatie van mitochondria en andere organellen, condensatie van chromosomen en microtubuli -assemblage, onder andere gebeurtenissen.
Mitoses fasen
Mitose begint met het einde van de G2 -fase en is verdeeld in vijf fasen: Profase, belofte, metafase, anafase en telofase. Ze gebeuren allemaal continu.
Kan u van dienst zijn: Cellulaire communicatie: typen, belang, voorbeeldenProfet
 Profet. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons
Profet. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons In dit stadium is de assemblage van de mitotische spil of mitotische apparaten het belangrijkste evenement. Profase begint met chromatine -verdichting en vormt chromosomen.
Elk chromosoom heeft een paar zusterschromaten, met identiek DNA, die nauw verbonden zijn in de buurt van hun centromeren. Eiwitcomplexen genaamd cohesines nemen deel aan deze unie.
Elke centromeer is gekoppeld aan een cinetocoro, een eiwitcomplex dat bindt aan microtubuli. Met deze microtubulo's kunnen elke kopie van de chromosomen worden toegewezen aan de dochtercellen. De microtubuli radia van elk uiteinde van de cel en vormen het mitotische apparaat.
In diercellen, vóór de profase, treedt de duplicatie van de centrering voor, het belangrijkste organiserende centrum van de microtubuli en de plaats waar de ouder- en zoon centriolen zich bevinden. Elke centrering bereikt de tegenoverliggende pool van de cel, waardoor een microtubulo -brug wordt opgezet tussen hen genaamd mitotisch apparaat.
In de meest recente evolutieplanten, in tegenstelling tot diercellen, zijn er geen centra en is de oorsprong van microtubuli niet duidelijk. In fotosynthetische cellen van oudere evolutionaire oorsprong, zoals groene algen, zijn er centra.
Belofte
Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] Mitose moet de segregatie van chromosomen en de verdeling van de nucleaire envelop van het nucleaire porie- en nucleoluscomplex garanderen. Afhankelijk van of de nucleaire wikkeling (in) verdwijnt of niet, en de mate van denstegratie van de in, varieert de mitose van gesloten tot volledig open.
Bijvoorbeeld in S. Cerevisae Mitose is gesloten, in NAAR. Nidulans Het is semi -open en bij mensen is het open.
Bij gesloten myitose bevinden de polaire lichamen van de spindel zich in de nucleaire envelop, die de nucleatiepunten vormen van nucleaire en cytoplasmatische microtubuli. Cytoplasmatische microtubuli interageren met de celcortex en met de chromosomen kneaders.
In de semi -absert -myitose, omdat de in gedeeltelijk desensamblada is, wordt de nucleaire ruimte binnengevallen door de microtubuli die uit de centra worden gekomen en door twee openingen van de in, gevormd gemaakt gemaakt door de in.
Bij open mitose komt de volledige de -slasamblage voor in, het mitotische apparaat is voltooid en chromosomen worden verplaatst naar het midden van de cel.
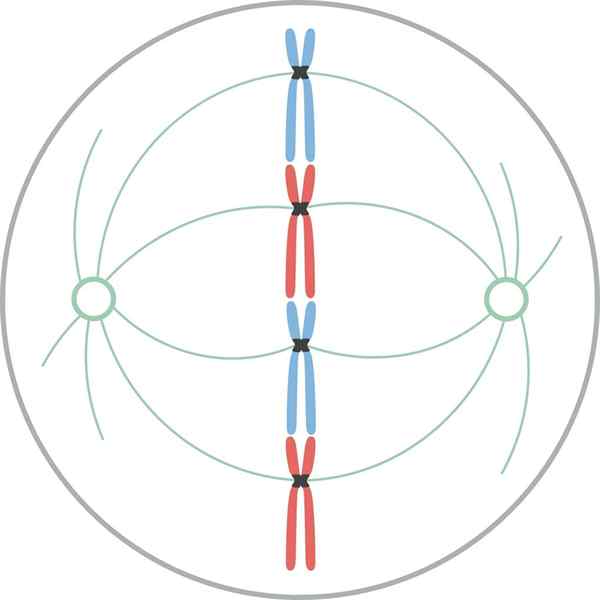
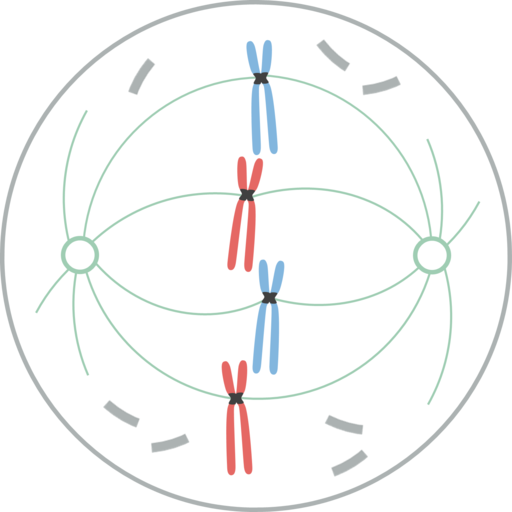
Metafase
Chromosomen uitgelijnd in de equatoriale plaat van de cel tijdens de mythische metafase In metafase zijn chromosomen uitgelijnd in de cel van de cel. Het denkbeeldige vlak loodrecht op de spindelas, die door de binnenomtrek van de cel gaat, wordt de metafase -schaal genoemd.
In zoogdiercellen is het mitotische apparaat georganiseerd in een centrale mitotische spil en een paar osteres. De mitotische spindel bestaat uit een symmetrische bilaterale bundel van microtubuli die is verdeeld in de cel van de cel, die twee tegenovergestelde helften vormt. De Ásteres bestaan uit een groep microtubuli in elke spindelpaal.
In het mitotische apparaat zijn er drie groepen microtubuli: 1) astrale, die de rster vormen, beginnen vanuit het centreren en stralen naar de celcortex; 2) van de cinetocoro, die binden aan de chromosomen door de cinetocoro; en 3) polair, die zich verbinden met de microtubuli van de tegenoverliggende pool.
In alle hierboven beschreven microtubuli zijn de uiteinden (-) gericht op de centra.
In plantencellen, als er geen centreer is, is de spil vergelijkbaar met die van diercellen. De spil bestaat uit twee helften met tegengestelde polariteit. De uiteinden (+) worden gevonden in de equatoriale plaat.
Het kan u van dienst zijn: podocyten: kenmerken, histologie en functiesAnafase
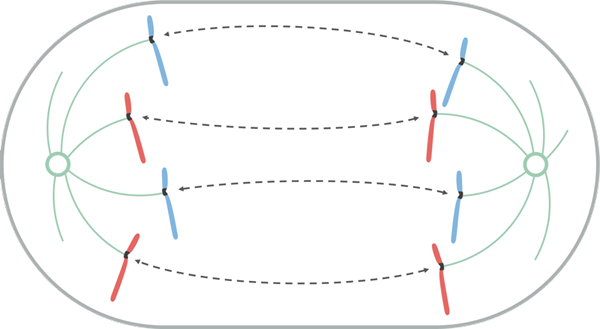 Bron: Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons
Bron: Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)], van Wikimedia Commons Anafase is vroeg en laat verdeeld. In vroege anafase vindt de scheiding van zusterchromatiden plaats.
Deze scheiding treedt op omdat de eiwitten die de unie behouden, zijn vastgebonden en omdat er een verkorting van de cynetocoro -microtubuli is. Wanneer het paar zusterchromatiden gescheiden zijn, worden ze chromosomen genoemd.
Tijdens de verplaatsing van de chromosomen naar de polen beweegt de cinetocoro langs de microtubule van dezelfde cynetocoro als zijn einde (+) dissocieert. Daarom is de beweging van chromosomen tijdens mitose een passief proces dat geen motorische eiwitten nodig heeft.
In de late anafase gebeurt een grotere scheiding van de polen. Een KRP -eiwit, gekoppeld aan het extreme (+) van de polaire microtubuli, in het overlappingsgebied van hetzelfde, marcheert tegen het einde (+) van een aangrenzende polaire microtubulus antiparallel. Aldus duwt de KRP de aangrenzende polaire microtubule tegen het einde (-).
In plantencellen, na de scheiding van chromosoom. Deze structuur maakt het begin mogelijk van het cytocinetische apparaat, genaamd Framoplasto.
Telofase
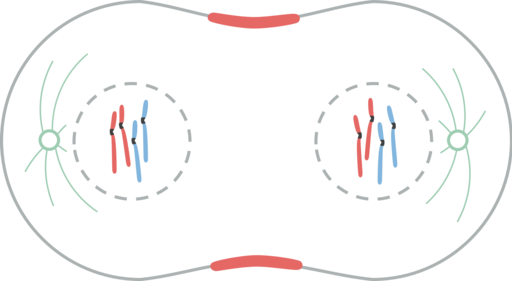 Telofase. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)]
Telofase. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] In de telofase gebeuren er verschillende gebeurtenissen. Chromosomen bereiken polen. Cinetocoro verdwijnt. Polaire microtubuli blijven zichzelf verlengen en bereiden de cel voor op cytokinese. De nucleaire envelop wordt opnieuw gevormd door fragmenten van de moeder om te wikkelen. De nucleolus verschijnt opnieuw. Chromosomen worden verkeerd begrepen.
Cytokinese

Citocinesis is de fase van de celcyclus waarin de cel is verdeeld. In diercellen vindt cytosinese plaats door middel van een actineflanstandsbouwriem. Deze filamenten glijden over elkaar, de diameter van de riem neemt af en een groef van clivaje wordt gevormd rond de celomtrek.
Omdat de vernauwing doorgaat, wordt de groef verdiept en wordt een intercellulaire brug gevormd, die het gemiddelde lichaam bevat. In het centrale gebied van de intercellulaire brug zijn de stralen van de microtubuli, die worden bedekt door een elektrodense matrix.
De breuk van de intercellulaire brug tussen post-mitotische zustercellen vindt plaats door abscissie. Er zijn drie soorten abscissie: 1) mechanisch breukmechanisme; 2) het indienen van mechanisme door interne blaasjes; 3) Plasmamembraan vernauwing voor splijting.
In plantencellen worden membraancomponenten binnen geassembleerd en wordt de celplak gevormd. Deze plaat groeit naar het oppervlak van het plasmamembraan, vermengt ermee en scheidt de cel in twee. Dan wordt de cellulose afgezet op het nieuwe plasmamembraan en vormt de nieuwe celwand.
Meiosis
Meiose is een soort celdeling die het aantal chromosomen in de helft vermindert. Aldus is een diploïde cel verdeeld in vier haploïde dochtercellen. Meiose gebeurt in kiemcellen en geeft aanleiding tot gameten.
De stadia van meiose bestaan uit twee divisies van de kern en cytoplasma, namelijk meiose I en Meiosis II. Tijdens meiose I scheiden de leden van elk paar homologe chromosomen. Tijdens meiose II scheiden zusterchromatiden en worden vier haploïde cellen geproduceerd.
Elke fase van mitose is verdeeld in profase, beloofd, metafase, anafase en telofase.
Kan u bedienen: Bacteriële celwand: kenmerken, biosynthese, functiesMeiosis I
- Profase I. De condensie van de chromosomen en de spil begint zich te vormen. DNA is verdubbeld. Elk chromosoom bestaat uit zusterchromatiden, samen met de centromere. Homologe chromosomen verschijnen tijdens synaps, waardoor cross -linking mogelijk is, wat de sleutel is tot het produceren van verschillende gameten.
- Metafase I. Het paar homologe chromosomen zijn uitgelijnd langs de metafaseplaat. Chiasma helpt het bijgevoegde paar te behouden. De microtubuli van de cinetocoro van elke pool sluiten zich aan bij een centromeer van een homoloog chromosoom.
- Anafase I. De microtubuli van de cinetocoro worden ingekort en de homologe paren zijn gescheiden. Een dubbele tegenhanger gaat naar een celpaal, terwijl de andere dubbele tegenhanger naar de andere kant van de paal gaat.
- Telofase i. Afzonderlijke homologen vormen een groep in elke celpool. De nucleaire envelop is weer. Citocinesis gebeurt. De resulterende cellen hebben de helft van het aantal originele celchromosomen.
Meiosis II
- PROFASE II. Een nieuwe spindel wordt gevormd in elke cel en het celmembraan verdwijnt.
- Metafase II. Spindelvorming is voltooid. De chromosomen hebben zusterschromatiden, samengevoegd in de centromere, uitgelijnd langs de metafaseplaat. De microtubuli van de cinetocoro die beginnen met tegengestelde polen binden aan de centromeren.
- Anafase II. De microtubers worden ingekort, de centromeren zijn verdeeld, de zusterchromatiden scheiden en gaan naar tegenovergestelde polen.
- Telofase II. De nucleaire wikkel rond vier groepen chromosomen worden gevormd: vier haploïde cellen worden gevormd.
Belang
Door sommige voorbeelden wordt het belang van de verschillende soorten celdeling geïllustreerd.
- Mitose. De celcyclus heeft onomkeerbare punten (DNA -replicatie, scheiding van zusterchromatiden) en controlepunten (G1/S). P53 -eiwit is de sleutel tot het G1 -controlepunt. Dit eiwit detecteert DNA -schade, stopt de celdeling en stimuleert de activiteit van enzymen die de schade herstellen.
In meer dan 50% van de menselijke kankers heeft het p53 -eiwit mutaties die zijn vermogen om specifieke DNA -sequenties in te stellen, annuleren. P53 -mutaties kunnen worden veroorzaakt door carcinogenen, zoals sigarettenrook benzopyreen.
- Meiosis. Wordt geassocieerd met seksuele reproductie. Vanuit evolutionair oogpunt wordt aangenomen dat seksuele reproductie ontstond als een proces om DNA te herstellen. De in een chromosoom geproduceerde schade kan dus worden gerepareerd op basis van de informatie van het homologe chromosoom.
Er wordt aangenomen dat de diploïde toestand voorbij was in oude organismen, maar dat het meer relevantie begon te zijn naarmate het genoom groter werd. In deze organismen heeft seksuele reproductie de complementatie, DNA -reparatie en genetische variatie.
Referenties
- Alberts, B., Johnson, a., Lewis, J., en kool. 2007. Biologie van de celmoleculair. Garland Science, New York.
- Bernstein, h., Byers, g.S., Michod, r.EN. 1981. Evolutie van seksuele reproductie: het belang van DNA -reparatie, complementatie en variatie. Amerikaanse naturalist, 117, 537-549.
- Lodish, h., Berk, een., Zipurski, s.L., Matsudaria, p., Baltimore, D., Darnell, J. 2003. Cellulaire en moleculaire biologie. Pan -American Medica -redactie, Buenos Aires.
- Raven, p.H., Johnson, g.B., Losos, J.B., Zanger, s.R. 2005 Biologie. Hoger onderwijs, Boston.
- Solomon, B.M., Berg, L.R., Martin, D.W. 2008. biologie. Thomson, VS.
- « Teporingo -kenmerken, het gevaar van uitsterven, habitat
- Ricinus Commune -kenmerken, habitat, toxiciteit, gebruik »