

Tonoplasto -kenmerken en functies
- 4012
- 1096
- Dr. Rickey Hudson
Toneplast Het is de term die in de biologie wordt gebruikt om de interne membranen van vacuolen in plantencellen te identificeren. De toon heeft een selectieve permeabiliteit en omsluit water, ionen en opgeloste stoffen in de vacuoles.
Er zijn grondige studies naar de moleculaire samenstelling van de toon, omdat de transporteiwitten in deze membranen de groei van planten reguleren, stress tot zoutgehalte en uitdroging en gevoeligheid voor pathogenen.
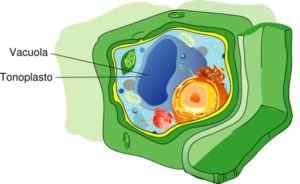 De toon van een plantencel (bron: Mariana Ruiz [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons)
De toon van een plantencel (bron: Mariana Ruiz [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons) Over het algemeen bevat de vacuole die de toon samenstelt, 57,2% van het gehele celvolume in planten. Dit percentage kan echter variëren, afhankelijk van de manier van leven, normaal gesproken de cactus- en woestijnplanten die minder of grotere grootte -vacuolen hebben.
In sommige soorten planten kan de vacuola gescheiden door de Toneplast tot 90% van het interieurvolume van alle plantencellen bezetten.
Omdat het betrokken is bij een constant verkeer van moleculen, ionen en enzymen tussen het cytosol en het interieur van de vacuola, is de toon rijk aan het transport van eiwitten, kanalen en aquaporines (poriën of kanalen waar het water passeert).
Veel van de interne blaasjes zoals fagosomen of transportblaasjes eindigen.
Biotechnologen richten hun inspanningen op de noodzakelijke technieken om, in planten van commerciële interesse, zoals tarwe en rijst, toon met de kenmerken van zoutoplossing -resistente planten op te nemen,.
[TOC]
Kenmerken
De toon bestaat meestal uit eiwitten en lipiden die worden besteld in de vorm van lipidebilay. In vergelijking met andere celmembranen heeft het echter unieke eiwitten en lipiden in zijn samenstelling.
Het vacuolaire membraan (de toon) bestaat uit 18% van neutrale lipiden en sterolen, 31% glycolipiden en 51% fosfolipiden. Normaal gesproken worden vetzuren aanwezig in lipiden die Bilay vormen.
Kan u van dienst zijn: diploïde cellenDe enorme vacuole gedefinieerd door de Toneplast begint als een set van meerdere kleine vacuolen die worden gesynthetiseerd in het endoplasmatisch reticulum, vervolgens zijn eiwitten uit het Golgi -apparaat opgenomen.
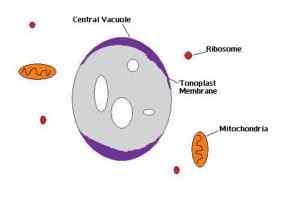 Schema van de centrale vacuola van een plantencel (Bron: I Am the Author: Gevictor [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons)
Schema van de centrale vacuola van een plantencel (Bron: I Am the Author: Gevictor [CC BY-SA 3.0 (https: // creativeCommons.Org/licenties/by-sa/3.0)] via Wikimedia Commons) Eiwitten van het Golgi -apparaat zijn kanalen, enzymen, transportbanden en structurele eiwitten en ankerglycoproteïnen die zich in de toon zullen positioneren.
Alle kleine vacuolen fuseren en organiseren langzaam en geleidelijk om de toon te vormen die aanleiding geeft tot een grote vacuola, vult voornamelijk water en ionen. Dit proces vindt plaats in alle organismen van het koninkrijk Plantae, Daarom hebben alle plantencellen een toon.
De toon, zoals de mitochondriale lipide bilay.
Functie
De hoofdfunctie van de toon is om te functioneren als een semipermeabele barrière, de ruimte afbakenen die door de vacuola wordt begrepen en het scheidt van de rest van het cytosolische gehalte.
Deze "semipermability" wordt benut door plantencellen voor turgiditeit, pH -controle, groei, naast vele andere functies.
Turgentie- en waterpotentieel
De meest bestudeerde functie van planten is het reguleren van celbegoedigheid. De concentratie ionen en water die zich binnen de vacuola bevinden, neemt deel, door het drukpotentiaal (ψp), in het waterpotentiaal (ψ) zodat de watermoleculen binnenkomen of in de cel vertrekken.
Dankzij de aanwezigheid van de toon wordt de drukpotentiaal (ψp) die de protoplast (plasmamembraan) op de celwand in de cellen uitoefent, gegenereerd. Deze kracht verwerft positieve waarden terwijl de vacuola druk uitoefent op de protoplast en dit op zijn beurt op de celwand.
Wanneer het water de vacuola door de toon verlaat en vervolgens de plantencel verlaat, begint de vacuola te samentrekken en gaat de turgiteit van de cel verloren, waardoor drukwaarden (ψp) dichtbij nul en zelfs negatief worden bereikt.
Kan u dienen: Sertoli -cel: kenmerken, histologie en functiesDit proces staat bekend als beginnende plasomolyse en dat is wat op zijn beurt de verwelking produceert die we in planten waarnemen.
Wanneer de plant wordt samengevoegd, neemt het osmotische potentieel (ψp) toe, omdat wanneer de concentratie kaliumionen (k+) in de cel groter is dan de concentratie van opgeloste stoffen buiten, het water naar binnen beweegt.
Deze kaliumionen (k+) bevinden zich meestal in de vacuola en zijn, toegevoegd met de cytosolionen, verantwoordelijk voor het genereren van het osmotische potentieel (ψp). De toon is permeabel voor deze kaliumionen dankzij een atpay die in zijn structuur heeft.
PH -onderhoud
Atasas in de toon.
Rootcelmembraan ATP's worden geactiveerd door de aanwezigheid van kaliumionen (k+), deze introduceren kalium (k+) ionen en verdrijven protonen (h+). Daarentegen worden de in de toon gevonden in de toon geactiveerd in aanwezigheid van chloor (Cl-) in de cytosol.
Deze regelen de concentratie van chloorionen (Cl-) en interne waterstof (H+). Beide ATP's werken in een soort "spel" om de pH in cytosol van plantencellen te regelen, om de pH te verhogen of te verlagen tot een pH van 7 of hoger in de cytosol.
Wanneer er een zeer hoge concentratie van protonen (H+) in het cytosol is, introduceert het celmembraan atpass kaliumionen (k+); Terwijl Atasa van de toon van chloorionen zuigt (Cl-) en waterstof (H+) van de cytosol in de binnenkant van de vacuola.
NAARionencumulatie
De tonoplast heeft verschillende soorten primaire protonenpompen. Bovendien heeft het transportkanalen voor calciumionen (Ca+), waterstofionen (H+) en andere ionen die specifiek zijn voor elke plantensoort.
Kan u van dienst zijn: exocytose: proces, typen, functies en voorbeeldenAtasas pompprotonen (H+) naar het interieur van de vacuola, waardoor het lumen hiervan een zure pH verwerft, met waarden tussen 2 en 5, en een positieve gedeeltelijke belasting. Deze pompen hydrolyseren de ATP in de cytosol en introduceren door een porie protonen (H+) in de richting van het lumen van de vacuola.
Pyrofosfase is een ander type "pompen" van de toon die ook protonen (H+) in de vacuole introduceren, maar ze doen het door de hydrolyse van het pyrofosfaat (PPI). Deze pomp is exclusief voor de planten en hangt af van de Mg ++ en K -ionen+.
In de tonoplast kunt u andere soorten ATASA's vinden die protonen naar het cytosol pompen en die calciumionen (Ca ++) in de binnenkant van de vacuola introduceren. Calcium (Ca ++) wordt gebruikt als een boodschapper in het cellulaire interieur en het lumen van de vacuola wordt gebruikt als een afzetting van deze ionen.
Misschien zijn de meest voorkomende eiwitten in de toon de calciumkanalen, deze maken calciumuitgang (Ca+) mogelijk geïntroduceerd door membraan Atasas.
Momenteel zijn ook primaire bommen of ABC -type transporters geïdentificeerd (uit het Engels NAARTP-BAansprakelijk CAsset) In staat om grote organische ionen in de vacuole te introduceren (zoals bijvoorbeeld glutathione).
Referenties
- Blumwald, E. (1987). Toneplast -blaasjes als hulpmiddel bij de studie van iontransport bij de plant -vacuole. Physiologia Plantarum, 69 (4), 731-734.
- Dean, J. V., Mohammed, l. NAAR., & Fitzpatrick, t. (2005). De vorming, vacuolaire lokalisatie en toupoplast van salicylzuur glucose -conjugaten in tabakscelophangculturen. Plant, 221 (2), 287-296.
- Gomez, L., & Chrispeels, m. J. (1993). Toneplast- en vacuolaire oplosbare eiwitten zijn het doelwit van verschillende mechanismen. De plantencel, 5 (9), 1113-1124.
- Jauh, g. EN., Phillips, T. EN., & Rogers, J. C. (1999). Tonoplast intrinsiek eiwit is vorm als markers voor vacuolaire functies. De plantencel, 11 (10), 1867-1882.
- Liu, l. H., Ludewig, u., Gassert, B., Uitmer, w. B., & von Wirén, n. (2003). Ureumtransport door stikstofgereguleerde tonoplast intrinsieke eiwitten in Arabidopsis. Plant Physiology, 133 (3), 1220-1228.
- Pesssarakli, m. (2014). Handboek van plant- en gewasfysiologie. CRC Press.
- Taiz, l., Zeiger, E., Møller, i. M., & Murphy, een. (2015). Plantenfysiologie en ontwikkeling