

Postmendelian erfgoed hoofdtheorieën

- 2482
- 264
- Aaron Okuneva
De Postmendeliaans erfgoed Het omvat alle ideeën en/of werken die zijn ontstaan na de studies die door de religieuze naturalist Gregorio Mendel in 1865 zijn gepubliceerd, en die genetische fenomenen of gedragingen verklaren die verschillen van die beschreven door dit.
Gregorio Mendel wordt beschouwd als de vader van de genetica, omdat het bestuderen van kruisen tussen erwtplanten, de eerste erfrechtwetten die meer dan 100 jaar geleden hebben opgezet. De eerste wet, "wet van segregatie" genoemd, stelt voor dat genen in verschillende of alternatieve vormen kunnen bestaan.
 Gregorio Mendel (Bron: Onbekende auteur / Public Domain, via Wikimedia Commons)
Gregorio Mendel (Bron: Onbekende auteur / Public Domain, via Wikimedia Commons) Deze alternatieve vormen worden "allelen" genoemd en beïnvloeden direct de fenotypische karakters van een soort. Allelen kunnen de kleur van het haar, de ogen, het type haar, enz. Bepalen.
In zijn eerste wet stelde Mendel voor dat elk fenotypische kenmerk van ouders op hun kinderen wordt bestuurd door twee allelen, elk van een ouder (een van de "moeder" en een van de "vader").
Volgens dit, wanneer de parentalen verschillende allelen leveren, resulteert de nakomelingen met een mengsel, waarbij een van de allelen dominant is (degene die tot expressie wordt gebracht in het fenotype) en de andere recessief is (het wordt niet gezien, of is niet 'latent ”).
De tweede wet van Mendel staat bekend als de "wet van onafhankelijke erfenis", omdat Mendel op zijn Cruces merkte dat het overervingspatroon dat de allelen kenmerkte die een kenmerk definieerde, geen invloed had op het overervingspatroon van de allelen die ze definieerden tegen een ander kenmerk.
[TOC]
Postmendeliaanse erfenistheorieën
Aangezien Gregor Mendel zijn beroemde wetten meer dan 100 jaar voorstelde, zijn nieuwe mechanismen opgehelderd om de erfenis van personages te verklaren, waarvan er veel van velen kon worden gezegd dat ze tegenspreken of aanzienlijk verschillen van die beschreven door Mendel.
Voorbeeld van de fenomenen die worden beschreven door de "moderne" wetenschap, wiens vooruitgang alleen mogelijk is geweest dankzij de voortgang van experimentele strategieën en onderzoeksmethoden, is de erfenis die verband houdt met seks, onvolledige dominantie en codominantie, onder andere.
- Erfenis gekoppeld aan seks
De erfenis die verband houdt met seks heeft te maken met de genen die alleen worden gevonden in sekschromosomen. Bij mensen wordt seks bepaald door de chromosomen x en y. Vrouwen hebben twee exemplaren van X (xx) en de mannen een van X en een andere van y (xy).
Kan u van dienst zijn: gekoppelde genen Overerving gekoppeld aan maternaal via (Bron: Jordi Picart/CC BY-SA (https: // creativeCommons.Org/licenties/by-sa/4.0) Via Wikimedia Commons)
Overerving gekoppeld aan maternaal via (Bron: Jordi Picart/CC BY-SA (https: // creativeCommons.Org/licenties/by-sa/4.0) Via Wikimedia Commons) De genen die in elk van deze twee chromosomen worden gevonden, zijn die welke bijdragen aan die eigenschappen die gekoppeld zijn aan seks. De genen op het X -chromosoom zijn aanwezig bij mannen en vrouwen, maar de genen van en zijn alleen bij mannen.
De erfenis die verband hield met seks werd waargenomen en voor het eerst beschreven door William Bateson, een van de eerste wetenschappers in het "herontdekken" van Mendel en profiteren van hen om nieuwe experimenten uit te voeren op basis van hun conclusies.
Bateson maakte experimentele crubs van kippen met plumages van verschillende kleuren, sommige met donkere en gladde veren en andere met gladde veren en met een gestreepte patroon.
Door deze kruisen merkte hij op dat in de nakomelingen de resulterende mannen altijd plumages hadden met een gestreepte patroon.
Alleen in de derde generatie, toen een mannetje krabde met een donkere en gladde veren gekruist, werden enkele hanen met zwart verenkleed verkregen. Dit aandeel als gevolg van de kruisen daagde de door Mendel voorgestelde verhoudingen uit.
Overerving gekoppeld aan X
Na het werk van Bateson vond Thomas Henry Morgan een logische verklaring voor dergelijke vreemde resultaten verkregen door Bateson, waarbij het kleurverschil in kippen toeschreef waaraan het gestreepte patroon werd gecodeerd door een dominant allel dat op het X -chromosoom stond.
In de kippen komt de heterogametische seks (XY) in tegenstelling tot de mens, ondertussen komt de homogamische seks overeen met de mannen (xx).
Het allel bestudeerd door Bateson en Morgan werd toen uitgedrukt door al die mannetjes met een gestreepte verenkleed, maar toen een man twee X -chromosomen erfde met het allel dat overeenkomt met de zwarte kleuring, verschenen mannen van donker verenkleed en zonder krassen patroon.
- Incomplete dominantie
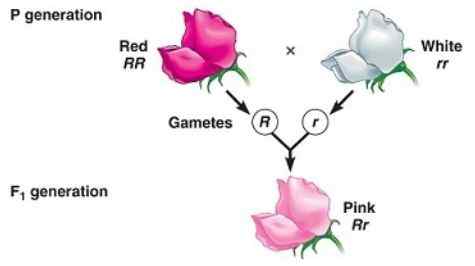 Tussentijdig fenotype in generatie F1 veroorzaakt door onvolledige dominantie
Tussentijdig fenotype in generatie F1 veroorzaakt door onvolledige dominantie Onvolledige dominantie is wanneer het fenotype dat wordt waargenomen in de nakomelingen dat het gevolg is van de kruising van twee homozygote individuen (de ene dominant en een ander recessief) een "tussenliggend" fenotype tussen het dominante en het recessieve.
Het heeft te maken met het "onvermogen" van het "dominante" allel van het volledig maskeren van de expressie van het recessieve allel, daarom komt het resulterende fenotype overeen met een mengsel van de kenmerken die worden gecodeerd door beide allelen, vaak beschreven als een "verdund fenotype "Van de dominante.
Kan u van dienst zijn: monosomieënBijvoorbeeld in de nachtelijke planten (Mirabilis Jalapa) De typische kleur van wilde bloemblaadjes is rood. Wanneer een plant met rode bloemblaadjes kruist met een pure lijnplant met witte bloemblaadjes, wordt verkregen dat al het nageslacht roze bloemblaadjes (roze).
- Codominantie
Wanneer twee dominante homozygote individuen worden gekruist en in de nakomelingen is er een derde fenotype dat overeenkomt met de twee kenmerken die worden bepaald door de twee ouderlijke allelen, praten we over codominance.
In dit geval, volgens een voorbeeld dat vergelijkbaar is met dat van de bloemen van het vorige voorbeeld, zou het "derde fenotype" niet overeenkomen met roze bloemen, maar met een plant met witte bloemen en rode bloemen in dezelfde stengel, sinds de twee allelen " Co -cake ”, en niemand maskeert de andere.
Vaak zijn echter onvolledige codominantie en dominantie moeilijk om fenotypisch te onderscheiden, omdat beide het eigen fenotype van een individu produceren dat twee verschillende allelen lijkt tot expressie te brengen.
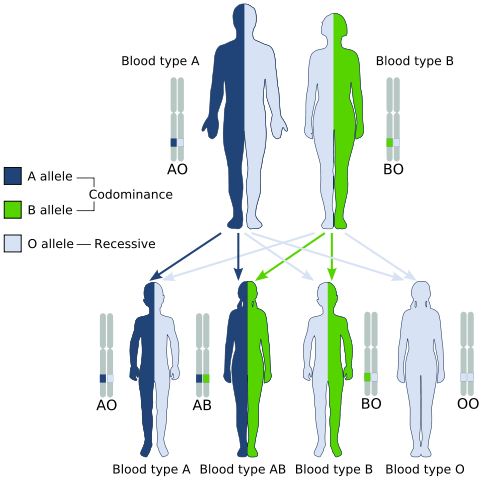 Codominance, voorbeeld van een type postmendeliaanse erfenis (Bron: GyssinemRabettalk✉ Deze W3C-RUN-gespecificeerde afbeelding werd gemaakt met Inkscape. / Public Domain, via Wikimedia Commons)
Codominance, voorbeeld van een type postmendeliaanse erfenis (Bron: GyssinemRabettalk✉ Deze W3C-RUN-gespecificeerde afbeelding werd gemaakt met Inkscape. / Public Domain, via Wikimedia Commons) Genotypisch aanwezig zijn echter individuen.
Een klassiek voorbeeld van codominantie is het type bloedgroep bij mensen, omdat allelen die coderen voor antigenen A en B in menselijke populaties codominant zijn.
Dat wil zeggen, wanneer allelen A en B samen worden gevonden in de cellen van een individu, heeft het een bloed -ab -type.
- Dodelijke allelen
Ze zijn recessieve allelen voor een bepaald karakter dat, wanneer ze homozygoot zijn, dodelijk zijn voor het individu. Personen die deze allelen presenteren, worden niet geboren, omdat ze het normale functioneren van moleculaire machines vermijden en vertaling van genen vermijden.
- Geninteracties
Veel plaatsen in het genoom van een individu interageren met andere delen van hetzelfde genoom, dat wil zeggen dat de punctuele modificatie van een allel in het genoom de expressie van andere allelen in de cellen van een individu direct kan conditioneren.
Het kan u van dienst zijn: sociale, economische en milieu -impact van genetische manipulatieDeze interacties zijn moeilijk te bepalen en worden ingedeeld in twee verschillende typen: de genen die op dezelfde genetische route interageren en die die dit doen op verschillende genetische routes.
Interacties op verschillende genetische routes
Een klassiek en goed bekend voorbeeld van genen die in verschillende genetische routes interageren, heeft te maken met de genen die de kleur van de huid van maïsslangen bepalen (Pantherophis guttatus), wiens gemeenschappelijke kleur een repetitief patroon is van zwarte en oranje plek.
 Corn Snake (Bron: Factumquintus, via Wikimedia Commons)
Corn Snake (Bron: Factumquintus, via Wikimedia Commons) Dit fenotype wordt geproduceerd door twee verschillende pigmenten die onder strikte genetische controle staan. Een gen bepaalt de sinaasappel en dit heeft twee allelen: een of+, die codeert voor het pigment en een andere O-, die de totale afwezigheid van het oranje pigment bepaalt.
Een ander ander gen bepaalt de aanwezigheid of afwezigheid van het zwarte pigment, de allelen van dit gen staan bekend als B+, degene die de aanwezigheid van het zwarte pigment en B- bepaalt, die de afwezigheid van het pigment bepaalt.
Beide genen zijn niet verbonden en onafhankelijk bepaald, de kleur van slangen, dat wil zeggen een slang met het genotype of+/B+ is een normale slang met oranje en zwart fenotype, maar elke genotypische variatie zal resulteren in een ander fenotype.
Interacties op dezelfde genetische route
Het klassieke voorbeeld van deze genen is wat is beschreven voor "Bells" -bloemen (uit de familie Campanulae), wiens "wilde" fenotype overeenkomt met planten met blauwe bloemen.
De blauwe kleur van de belbloemen wordt niet bepaald door een gen, maar door een metabole route waar verschillende enzymen, elk gecodeerd door een ander gen, deelnemen aan de synthese van een pigment.
Er zijn dus fenotypes die worden gekenmerkt door het resultaat te zijn van de interactie van verschillende genen, waarvan de producten deelnemen aan dezelfde metabole route en waar de mutatie of slechte functie van een van deze een fenotype oplevert, in dit geval van witte bloemen, afgewezen.
Referenties
- Ceccalli, D. (2019). Geschiedenis van erfenis.
- Griffiths, p., & Stotz, K. (2013). Genetica en filosofie: een inleiding. Cambridge University Press.
- Lewin, B., Krebs, J. EN., Goldstein, E. S., & Kilpatrick, s. T. (2014). Lewin's genen xi. Jones & Bartlett Publishers.
- McCleran, G. EN., Vogler, g. P., & Plomin, r. (1996). Genetica en gedragsmedicine. Gedragsmedicijn, 22(3), 93-102.
- Richmond, m. L. (2008). Het pre- en post-Mendeliaanse onderzoeksprogramma van William Bateson in 'Erfelijkheid en ontwikkeling.". Een culturele geschiedenis van erfelijkheid IV: erfelijkheid in de eeuw van het gen, 343, 213-242.
- « Cellulaire prikkelbaarheidsprincipes, componenten, antwoorden (voorbeelden)
- Uitgestorven en huidige megafauna (met voorbeelden) »