

Hollandse erfeniskenmerken, functies van genen, degeneratie
- 1057
- 149
- Kurt Aufderhar Jr.
De Hollandische erfenis Het is de overdracht van genen gekoppeld aan het seksuele chromosoom en van ouders naar kinderen. Deze genen worden intact overgedragen of geërfd, dat wil zeggen dat ze geen recombinatie lijden, zodat ze kunnen worden beschouwd als een enkel haplotype.
Het chromosoom en is een van de twee geslachtschromosomen die het biologische geslacht van het embryo bij mensen en andere dieren bepaalt. De vrouwtjes hebben twee X -chromosomen, terwijl de mannen een X -chromosoom en een chromosoom hebben en.
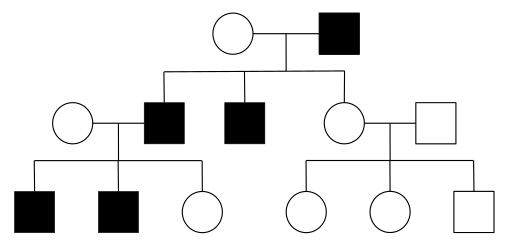
 Schema van het Hollandic Inheritance Patroon (Bron: MadIBC68 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons)
Schema van het Hollandic Inheritance Patroon (Bron: MadIBC68 [CC BY-SA 4.0 (https: // creativeCommons.Org/licenties/by-sa/4.0)] via Wikimedia Commons) De vrouwelijke gamete verzendt altijd een X -chromosoom, terwijl mannelijke gameten een X -chromosoom of chromosoom kunnen verzenden en daarom wordt gezegd dat "ze seks bepalen".
Als de vader een X -chromosoom overbrengt, zal het embryo genetisch vrouwelijk zijn, maar als de vader een chromosoom overbrengt en het embryo genetisch mannelijk zal zijn.
In het proces van seksuele reproductie worden de twee geslachtschromosomen gerecombineerd (ze wisselen genetische informatie met elkaar uit) die de functies combineren die door beide ouders worden overgedragen. Deze combinatie helpt mogelijke defecte kenmerken in het nageslacht te elimineren.
95 % van het chromosoom en is echter exclusief voor mannelijke organismen. Deze regio staat algemeen bekend als "specifieke mannelijke regio van Y", en het is niet seksueel recombineren met het X -chromosoom tijdens de reproductie.
Bovendien recombineren de meeste genen op het chromosoom en recombineren niet met enig ander chromosoom tijdens seksuele reproductie, omdat deze aan elkaar zijn gekoppeld, dus de meeste zijn hetzelfde bij ouderlijke en afstammelingen.
[TOC]
Chromosoom en
Het chromosoom en is de kleinste van alle chromosomen. Bij zoogdieren bestaat dit uit ongeveer 60 megabases en heeft slechts enkele genen. Het beschikbare gebied om te transcriberen (euchromatine) is 178 drieling, en de rest zijn pseudogenes of herhaalde genen.
Herhaalde genen worden gevonden in meerdere kopieën en palindromisch, wat betekent dat ze op dezelfde manier worden gelezen in beide richtingen zoals het woord "zwemmen"; Een DNA Palindrome -sequentie zou zoiets zijn als: Ataata.
Kan u van dienst zijn: wat is chromosomale permutatie? Human Chromosomes (Bron: National Center for Biotechnology Information, U.S. National Library of Medicine [Public Domain] via Wikimedia Commons)
Human Chromosomes (Bron: National Center for Biotechnology Information, U.S. National Library of Medicine [Public Domain] via Wikimedia Commons) Van de 178 eenheden of drieling worden blootgesteld aan transcriptie, 45 unieke eiwitten van dit chromosoom worden verkregen. Sommige van deze eiwitten worden geassocieerd met het geslacht en de vruchtbaarheid van het individu en andere niet -reproductieve zijn ribosomale eiwitten, transcriptiefactoren, enz.
De chromosoomarchitectuur is verdeeld in twee verschillende regio's, een korte arm (P) en een lange arm (Q). De korte arm bevat 10 tot 20 verschillende genen, omvat ongeveer 5 % van het gehele chromosoom en kan recombineren met het X -chromosoom tijdens meiose.
 Chromosoom en mensen. De kleine arm (P) en de grote (Q) (Bron: John W is geïdentificeerd. Kimball [CC door 3.0 (https: // creativeCommons.Org/licenties/door/3.0)] via Wikimedia Commons)
Chromosoom en mensen. De kleine arm (P) en de grote (Q) (Bron: John W is geïdentificeerd. Kimball [CC door 3.0 (https: // creativeCommons.Org/licenties/door/3.0)] via Wikimedia Commons) De lange arm vormt ongeveer de resterende 95% van het chromosoom en. Deze regio staat bekend als "niet -recombinant gebied" (Nry), hoewel sommige onderzoekers suggereren dat in die regio recombinatie optrad en de regio "specifiek mannelijk gebied" (RMS) (RMS) moet worden genoemd).
De genen die behoren tot het niet -recombinante gebied van Y (95%) hebben een Holland -erfenis, omdat ze zich uitsluitend in genoemde chromosoom bevinden en verenigd zijn of aan elkaar verbonden zijn tussen hen. Er is geen recombinatie in deze regio en de mutatiesnelheid is erg laag.
Functies van genen met hollandse erfenis
In 1905 merkten Nettie Stevens en Edmund Wilson voor het eerst op dat de cellen van mannen en vrouwen een andere chromosomale structuur hadden.
De vrouwencellen hadden twee kopieën van de grote chromosoom X, ondertussen hadden de mannen alleen een kopie van deze X en, geassocieerd met dit chromosoom, hadden een veel kleiner chromosoom, het chromosoom en.
In de eerste 6 weken van de zwangerschap ontwikkelen alle embryo's, hetzij genetisch vrouwelijk of mannelijk, op dezelfde manier. In feite, als ze het tot de bevalling zouden blijven doen, zouden ze aanleiding geven tot een fysiek vrouwelijke pasgeborene.
Kan u van dienst zijn: cytoplasmatisch erfgoedAl deze veranderingen in mannelijke embryo's voor de werking van het gen genaamd "Seksuele bepaling regio en" gelegen op het chromosoom en. Dit is afgeleid van het Engels "Seksbepalende regio en'En hij afkortte literatuur zoals Sry.
Het Sry-gen werd in 1990 ontdekt door Robin Lovell-Badge en Peter Goodfellow. Alle embryo's met een actieve kopie van dit gen ontwikkelen penis, testikels en baard (op volwassen leeftijd).
Dit gen werkt als een schakelaar. Wanneer "ontsteken" wordt geactiveerd door mannelijkheid en wanneer het "uit" is, geeft het aanleiding tot vrouwelijke individuen. Het is het meest bestudeerde gen van het chromosoom en reguleert vele andere genen die verband houden met het geslacht van individuen.
Het SOX9 -gen codeert voor een transcriptiefactor die cruciaal is bij de vorming van de testikels en wordt tot expressie gebracht in combinatie met het SRY -gen. Het SRY -gen activeert de expressie van SOX9 om de ontwikkeling van mannelijke gonaden bij veel dieren te initiëren.
Degeneratie van genen met hollandse erfenis
Alle genen gevonden in het chromosoom en, inclusief die welke worden overgedragen door nederlandse overerving, worden gevonden in een dwergchromosoom. Terwijl het X -chromosoom meer dan 1000 genen heeft, het chromosoom en minder dan 100 heeft.
Het chromosoom en was ooit identiek van grootte van het X -chromosoom. Gedurende ongeveer 300 miljoen jaar is de omvang echter geleidelijk afgenomen, tot het punt dat het minder genetische informatie heeft dan enig ander chromosoom.
Bovendien heeft het X -chromosoom een tegenhanger, omdat het bij vrouwen in paren (xx) maar het chromosoom verschijnt en alleen bij mannen wordt gevonden en geen tegenhanger heeft. De afwezigheid van een paar voorkomt het chromosoom en het recombineren van alle delen met een gelijke.
Deze afwezigheid van een paar voorkomt genen met een nederlandse erfenis, exclusief het chromosoom en kan zichzelf beschermen tegen mutaties en normale genetische achteruitgang van nucleïnezuren.
Kan u van dienst zijn: pleiotropyDe afwezigheid van recombinatie betekent dat elke mutatie die optreedt in de genen die gekoppeld zijn aan het chromosoom en of met een Holland -overerving intact wordt overgedragen aan de afstammelingen van mannelijk geslacht, wat een groot nadeel kan betekenen.
Hoewel het chromosoom en hun genen gedegenereerd zijn en kwetsbaar zijn voor mutaties, denken wetenschappers dat het verre van zichzelf beschadigt of verdwijnt, omdat sommige genen van dit chromosoom belangrijk zijn voor de spermaproductie.
De spontane mutaties die beschadigen of inactiveren zijn betrokken bij de productie van sperma, worden "geselecteerd", waardoor de vruchtbaarheid van de ouder met die mutatie wordt verminderd, waardoor dit zijn genen naar de nakomelingen wordt voorkomen.
Referenties
- Bradbury, n. NAAR. (2017). Alle cellen hebben een geslacht: studies van geslachtschromosoomfunctie op cellulair niveau. In principes van genderspecifieke geneeskunde (PP. 269-290). Academische pers.
- Buchen, l. (2010). De wispelturige en chromosoom.
- Carvalho, een. B., Dobo, b. NAAR., Vibranovski, m. D., & Clark, tot. G. (2001). Identificatie van vijf nieuwe genen op het en chromosoom van Drosophila melanogaster. Proceedings of the National Academy of Sciences, 98 (23), 13225-13230.
- Charlesworth, B., & Charlesworth, D. (2000). De degeneratie van en chromosomen. Filosofische transacties van de Royal Society of London. Serie B: Biological Sciences, 355 (1403), 1563-1572.
- Colaco, s., & Modi, D. (2018). Genetica van het mens en chromosoom en de associatie met mannelijke invertiliteit. Reproductieve biologie en endocrinologie, 16 (1), 14.
- Gerrard, D. T., & Filatov, D. NAAR. (2005). Positieve en negatieve selectie op zoogdier en chromosomen. Molecular Biology and Evolution, 22 (6), 1423-1432.
- Hughes, J. F., Skaletsky, h., Pyntikova, t., Minx, p. J., Serieus, T., Rozen, s. & Page, D. C. (2005). Behoud van Y-gekoppeld tijdens de menselijke evolutie onthult door vergelijkende sequencing in chimpanseee. Natuur, 437 (7055), 100.
- Komori, s., Kato, h., Kobayashi, s. Je., Koyama, k., & Isojima, s. (2002). Transmissie van en chromosomale microdeleties van vader op zoon Throuch intracytoplasmatische sperma -injectie. Journal of Human Genetics, 47 (9), 465-468.
- Malone, J. H. (2015). Widepraad redding van Y-gekoppelde genen door genement voor autosomen. Genome Biology, 16 (1), 121.
- Papadopulos, a. S., Chester, M., Ridout, k., & Filatov, D. NAAR. (2015). Snelle en degeneratie- en doseringscompensatie in plantensekschromosomen. Proceedings of the National Academy of Sciences, 112 (42), 13021-13026.
- « Mitochondriale erfenisstoepassingen, pathologieën, voorzorgsmaatregelen
- HapLootype Study -methoden, diagnostiek, ziekten »