

Donkere fase van fotosynthese
- 4252
- 1202
- Cecil Graham
De donkere fase van fotosynthese is het biochemische proces waardoor planten koolstofdioxide omzetten in glucose Wat is de donkere fase van fotosynthese?
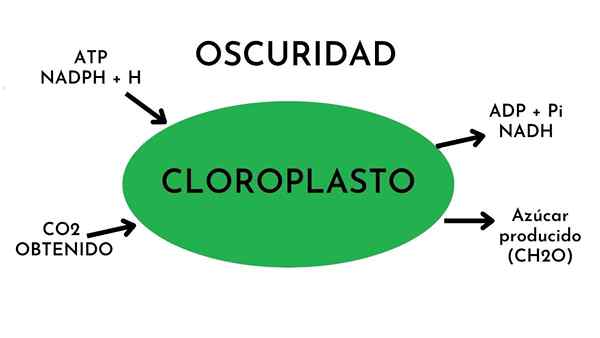
De Donkere fase van fotosynthese Het is het biochemische proces waardoor planten koolstofdioxide omzetten in glucose. Er wordt aan het donker verteld omdat licht niet nodig is voor het proces. Het staat ook bekend als koolstoffixatiefase of Calvin-Benson-cyclus. Dit proces vindt plaats in de Chloroplast stroma.
In de donkere fase wordt chemische energie geleverd door producten die in de lichtfase worden gegenereerd. Deze producten zijn de ATP -energiemoleculen (adenosine -trfosfaat) en de NADPH (een verminderde elektronendrager).
De fundamentele grondstof voor het proces in de donkere fase is koolstof, die wordt verkregen uit koolstofdioxide. Het eindproduct is koolhydraten of eenvoudige suikers.
Deze verkregen koolstofverbindingen zijn de fundamentele basis van de organische structuren van levende wezens.
Kenmerken van de donkere fase van fotosynthese
- Het wordt donker genoemd door geen directe deelname van zonlicht te vereisen voor de ontwikkeling ervan. Deze cyclus vindt plaats gedurende de dag of nacht.
- De donkere fase ontwikkelt zich voornamelijk in het stroma van chloroplast in de meeste fotosynthetische organismen. Het stroma is de matrix die de interne holte van de chloroplast rond het tilacoïde systeem vult (waar de lichtfase wordt uitgevoerd).
- In het stroma zijn de enzymen die nodig zijn om de donkere fase op te treden. De belangrijkste van deze enzymen is Rubisco (ribulous bifosfaatcarboxylase/oxygenase), het meest voorkomende eiwit, dat tussen 20 en 40% van alle bestaande oplosbare eiwitten vertegenwoordigt.
Mechanismen
De koolstof die nodig is voor het proces is in de vorm van co₂ (koolstofdioxide) in de omgeving. In het geval van algen en cyanobacteriën wordt de co₂ opgelost in het omringende water. In het geval van planten bereikt CO ₂ fotosynthetische cellen door huidmondjes (epidermale cellen).
Kan u van dienst zijn: AshCalvin-Benson-cyclus
Deze cyclus heeft verschillende reacties:
Eerste reactie
De co₂ Het kijkt naar een acceptorverbinding van vijf koolstofatomen (ribulosa 1,5-biphosfaat of rubp). Dit proces wordt gekatalyseerd door het Rubisco -enzym. De resulterende verbinding is een molecuul met zes koolstofarme.
Het breekt snel en vormt twee verbindingen van elk drie koolstofatomen (3-fosfoglycerate of 3pg).
Tweede proces
In deze reacties wordt de energie gebruikt door de ATP uit de lichtfase. Een fosforylering bevorderd door ATP -energie en een proces van een reductie gemedieerd door NADPH vindt plaats. Aldus wordt 3-fosfoglyceraat gereduceerd tot glyceraldehyde 3-fosfaat (G3P).
De G3P is een fospatada -suiker met drie koolstofarme, ook wel Triosa -fosfaat genoemd. Slechts een zesde deel van het 3-fosfaatglyceraldehyde (G3P) wordt omgezet in suikers als een product van de cyclus.
Dit fotosynthetische metabolisme wordt C3 genoemd, omdat het verkregen basisproduct een drie -koolstofuiker is.
Eindproces
De G3P -onderdelen die niet worden omgezet in suikers, worden verwerkt om monofosfaatribulous (Rump) te vormen. Rump is een tussenproduct dat transformeert in ribulous van 1,5 biphosfaat (rubp). Op deze manier wordt de CO₂ -acceptor hersteld En de Kelvin-Benson-cyclus sluit.
Van de totale RUBP geproduceerd in de cyclus op een typisch blad, wordt slechts één derde een zetmeel. Dit polysacharide wordt opgeslagen in chloroplast als een bron van glucose.
Een ander deel wordt omgezet in sucrose (een disaccharide) en getransporteerd naar andere organen van de fabriek. Vervolgens wordt sucrose gehydrolyseerd om monosachariden te vormen (glucose en vruchtbaar).
Andere fotosynthetische metabolismen
In het bijzonder omgevingscondities is het fotosynthetische plantenproces geëvolueerd en is efficiënter geworden. Dit heeft geleid tot het uiterlijk van verschillende metabole routes om suikers te verkrijgen.
Kan je dienen: Western Tuja: kenmerken, habitat, homeopathie, teeltC4 -metabolisme
In warme omgevingen zijn de bladstroom overdag gesloten om het verlies van waterdamp te voorkomen. Daarom neemt de concentratie van CO₂ in het blad af in relatie tot zuurstof (of2)). Het enzym Rubisco heeft een dubbele affiniteit van substraat: co₂ en o2.
Bij lage concentraties van co₂ en hoog2, Rubisco katalyseert de condensatie van O2. Dit proces wordt fotorerderspiratie genoemd en vermindert de fotosynthetische efficiëntie. Om fotoreerspiratie tegen te gaan, hebben sommige tropische omgevingen een bepaalde fotosynthetische anatomie en fysiologie ontwikkeld.
Tijdens het C4-metabolisme wordt koolstof gefixeerd in de mesofylcellen en treedt de Calvin-Benson-cyclus op in chlorofylische mantelcellen. De fixatie van de co₂ vindt 's nachts plaats. Het gebeurt niet in het stroma van de chloroplast, maar in de cytosol van de mesofylcellen.
De fixatie van de CO₂ vindt plaats door een carboxyleringsreactie. Het enzym dat de reactie katalyseert, is fosfoenolpyruvaatcarboxylase (pep-carboxylase), dat niet gevoelig is voor lage concentraties van co₂ In de cel.
Het co -acceptor molecuul is fosfoenolpirúviczuur (PEPA). Het verkregen tussenliggende product is oxaloazinezuur of oxalacetaat. Oxalacetaat wordt in sommige soorten planten of aspartaat (een aminozuur) gereduceerd tot malato.
Vervolgens gaat het kwaad naar de vasculaire fotosynthetische podcellen. Hier is het gedecarboxyleerd en pyruvaat en co₂ treedt op.
De co₂ komt de Calvin-Benson-cyclus binnen en reageert met de Rubisco om PGA te vormen. Van zijn kant keert het pyruvaat terug naar mesofylcellen, waar het reageert met ATP om de carbon -dioxide -acceptor te regenereren.
CAM -metabolisme
Het zure metabolisme van de crrasulaceae (CAM) is een andere strategie voor de fixatie van de co₂. Dit mechanisme is onafhankelijk geëvolueerd in verschillende groepen sappige planten.
Het kan je van dienst zijn: dicotyledonousCAM -planten gebruiken zowel de C3- als de C4 -weg, zoals in C4 -planten. Maar de scheiding van beide metabolismen is tijdelijk.
De co₂ wordt 's nachts ingesteld door de activiteit van het pep-carboxylase in het cytosol en vormt oxalacetaat. Oxalacetaat wordt gereduceerd tot malato, dat wordt opgeslagen in de vacuola als een formeel zuur.
Vervolgens wordt in aanwezigheid van het licht het lastuagezuur teruggewonnen uit de vacuola. Het is gedecarboxyleerd en de co₂ wordt overgebracht naar de rubp van de Calvin-Benson-cyclus in dezelfde cel.
CAM -planten hebben fotosynthetische cellen met grote vacuolen waar ricaanzuur wordt opgeslagen en chloroplasten waarbij de co₂ -geobgenoemde uit het muzikale zuur wordt omgezet in koolhydraten.
Eindproducten
Aan het einde van de donkere fase van fotosynthese worden verschillende suikers geproduceerd. Sucrose is een tussenproduct dat snel wordt gemobiliseerd van de bladeren naar andere delen van de plant. Het kan direct worden gebruikt om glucose te verkrijgen.
Zetmeel wordt gebruikt als reserve -stof. Het kan zich op het vel ophopen of worden getransporteerd naar andere organen, zoals stengels en wortels. Er wordt gehandhaafd totdat het vereist is in verschillende delen van de plant. Het wordt opgeslagen in speciale plastiden, genaamd Amyloplast.
De producten verkregen uit deze biochemische cyclus zijn van vitaal belang voor de plant. De geproduceerde glucose wordt gebruikt als koolstofbron om verbindingen vast te stellen, zoals aminozuren, lipiden of nucleïnezuren.
Aan de andere kant vertegenwoordigt het suikersproduct van de gegenereerde donkere fase de basis van de voedselketen. Deze verbindingen vertegenwoordigen zonne -energiepakketten omgezet in chemische energie, gebruikt door alle levende organismen.
Referenties
- Raven, p.H., R.F. Evert en S.EN. Eichhorn (1999). Biologie van planten. WH Freeman en Company Worth Publishers.
- Solomon, E.P., L.R. Berg en D.W. Martin (2001). biologie. McGraw-Hill Inter-American.