

Transcriptiefactormechanisme van werking, typen, functies
- 1658
- 235
- Glen Armstrong
A Transcriptiefactor Het is een regulerend "accessoire" eiwit dat nodig is voor genetische transcriptie. Transcriptie is de eerste stap van genetische expressie en impliceert de overdracht van informatie in het DNA naar een RNA -molecuul, dat vervolgens wordt verwerkt om aanleiding te geven tot genproducten.
RNA -polymerase II is het enzym dat wordt beschuldigd van de transcriptie van de meeste eukaryotische genen en produceert, naast enkele kleine RNA's, de messenger -RNA's die vervolgens worden vertaald in eiwitten. Dit enzym vereist de aanwezigheid van een type transcriptiefactoren die bekend staan als algemene of basale transcriptiefactoren.
Dit zijn echter niet de enige transcriptfactoren die in de natuur bestaan, omdat er "niet-generaal" eiwitten zijn, zowel in eukaryoten als in prokaryoten en bogen, die betrokken zijn bij de regulatie van de weefselspecifieke genetische transcriptie (in meercellige organismen ) of in de regulatie van genactiviteit in reactie op verschillende stimuli.
Deze transcriptiefactoren zijn effectoren van groot belang en kunnen vrijwel in alle levende organismen worden gevonden, omdat ze de belangrijkste bron van regulatie van genetische expressie vertegenwoordigen.
Gedetailleerde studies van verschillende transcriptiefactoren in verschillende soorten levende organismen geven aan dat ze een modulaire structuur hebben, waarbij een specifiek gebied verantwoordelijk is voor interactie met DNA, terwijl anderen de stimulerende of remmende effecten produceren.
De transcriptiefactoren nemen dan deel aan de modellering van de genetische expressiepatronen die niets te maken hebben met de veranderingen in de DNA -sequentie, maar met epigenetische veranderingen. De wetenschap die verantwoordelijk is voor de studie van deze veranderingen staat bekend als epigenetica.
[TOC]
Werkingsmechanisme
Om hun functies uit te oefenen, moeten transcriptiefactoren in staat zijn om een bepaalde DNA -sequentie te herkennen en specifiek samen te voegen om de transcriptie van dit gebied van het DNA positief of negatief te beïnvloeden.
De algemene transcriptiefactoren, die in principe hetzelfde zijn voor de transcriptie van alle type II -genen in eukaryoten, worden eerst geassembleerd op het promotorgebied van het gen, waardoor de positionering van het polymerase -enzym en de "opening" van de dubbele propeller wordt gericht.
Het proces wordt gegeven door verschillende opeenvolgende stappen:
- Unie van de algemene transcriptiefactor tfiid tot een reeks herhaalde timina (t) en adenine (a) in het gen dat bekend staat als "de doos Tata”; Dit veroorzaakt DNA -vervorming die nodig is voor de vereniging van andere eiwitten naar het promotoregebied.
- Achterste assemblage van andere algemene factoren (tfiib, tfiih, tfih, tfiie, tfiif, etc.) en van de RNA -polymerase II, die vormt wat de naam wordt genoemd Transcriptie -initiatiecomplex.
Kan u van dienst zijn: chromatiden- Bevrijding van het initiatiecomplex, fosforylering van polymerase door de TFIIH -factor en het begin van de transcriptie en synthese van een RNA -molecuul uit de sequentie van het gen dat wordt getranscribeerd.
Activering en repressie van transcriptie
Zoals vermeld, kunnen "niet -generale" transcriptiefactoren de expressie van genen reguleren, hetzij positief of negatief.
Activering
Sommige van deze eiwitten bevatten, naast de structurele domeinen van DNA -unie, andere redenen die bekend staan als activeringsdomeinen, die rijk zijn aan zuurzuurresten, glutamine of prolineresten.
Deze activeringsdomeinen werken samen met de elementen van de algemene transcriptiefactoren complex of met gerelateerde coactiverende moleculen die direct interageren met het complex. Deze interactie resulteert in de stimulatie van de assemblage van het transcriptieve complex of in de toename van zijn activiteit.
Repressie
De meeste transcriptiefactoren remmen de transcriptie bij het verstoren van de activiteit van transcriptiefactoren die positief werken, waardoor het stimulerende effect wordt geblokkeerd. Ze kunnen werken door de unie van de positieve factor voor het DNA te blokkeren of te werken op de factoren die de structuur van de chromatine inactiveren.
Andere remmende factoren werken direct de transcriptie blokkeren, zonder de werking van een activerende transcriptionele factor te blokkeren; en het basale transcriptieniveau verminderen, tot een nog lager niveau dan dat wordt bereikt in afwezigheid van activerende factoren.
Net als het activeren van eiwitten, werken onderdrukfactoren direct of indirect met basale of algemene transcriptiefactoren.
Jongens
Hoewel de meeste transcriptiefactoren worden geclassificeerd volgens de kenmerken of identiteit van hun DNA -vakbondsdomeinen, zijn er sommigen, ook geclassificeerd als transcriptiefactoren, die niet rechtstreeks interageren met DNA en bekend staan als transcriptiefactoren "indirect".
Directe transcriptiefactoren
Het zijn de meest voorkomende transcriptiefactoren. Ze hebben DNA -vakbondsdomeinen en kunnen genexpressie activeren of remmen door middel van hun unie naar specifieke DNA -regio's. Ze verschillen van elkaar, vooral met betrekking tot hun DNA -vakbondsdomeinen en hun oligomerisatietoestand.
De meest bestudeerde en erkende families van dit soort factoren zijn:
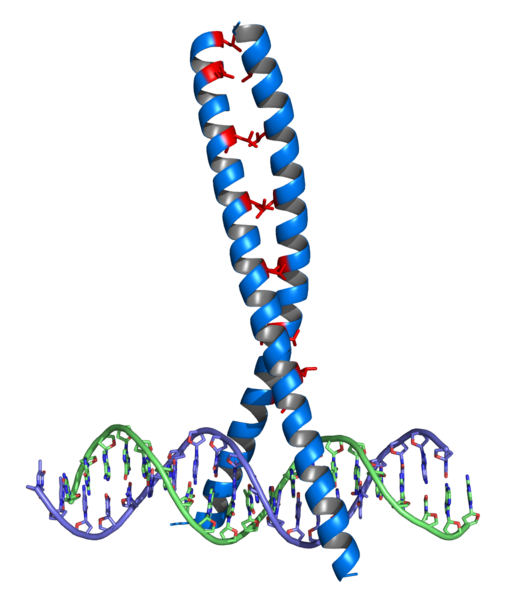
Helix-Gutero-Hélice ("Helix-turn-helix”, Hth)
Dit was de eerste familie van factoren met DNA -vakbondsdomeinen die werden ontdekt, en is aanwezig in veel eukaryoten en prokaryoten. De reden voor herkenning bestaat uit een α -propeller, een beurt en een tweede propeller α.
Ze hebben glycine domeinen in het gebied van de beurt behouden en ook wat hydrofoob afval dat helpt de opstelling van de twee propellers in de HTH -eenheid te stabiliseren.
Kan u van dienst zijn: genetische poolHomeodominium
Het is aanwezig in een groot aantal eukaryotische regulerende eiwitten. De eerste sequenties werden erkend in regulerende eiwitten van de ontwikkeling van Drosophila. Dit domein bevat een HTH-reden om lid te worden van het DNA en een extra α-propeller, naast een uitgebreide N-terminale arm.
Zinkvingers
Werden ontdekt in de transcriptiefactor tfiiia van Xenopus En er is aangetoond dat ze deelnemen aan vele aspecten van eukaryotische genetische regulatie. Ze worden gevonden in eiwitten geïnduceerd door signalen van differentiatie en groei, in protooncogenes en in sommige algemene transcriptiefactoren.
Ze worden gekenmerkt door de aanwezigheid van herhalingen in een partij van de motieven van zink van 30 afval die meerdere afval- en histidine -afval bevatten.
Steroïde receptoren
Deze familie omvat belangrijke regulerende eiwitten die, naast het hebben van een domein voor de Unie van hormonen, een domein van DNA -unie hebben en meestal fungeren als transcriptionele activatoren.
Union -domeinen bevatten 70 afval, waaronder 8 behouden cysteïneresten behouden. Sommige auteurs zijn van mening dat deze factoren een paar zinkvingers kunnen vormen, gezien de aanwezigheid van twee vier -cystein -spellen.
Leucina en helik-bucle-harice sluiting ("Helix-lus-helix "))
Deze transcriptiefactoren zijn betrokken bij differentiatie en ontwikkeling en werken voor de vorming van een heterodimeer. Het Leucin -sluitingsdomein wordt waargenomen in verschillende eukaryoten en wordt gekenmerkt door twee subdomeinen: de sluiting van leucines die dimerisatie meten en een basisgebied voor vereniging met DNA.
Redenen β Redenen
Ze worden voornamelijk gevonden in eukaryotische factoren en Distingu.
Indirecte transcriptiefactoren
Dit type transcriptiefactoren oefent zijn regulerende effecten op genetische expressie uit, niet door zijn directe interactie met DNA, maar door eiwit-eiwit interacties met andere transcriptiefactoren die welwerken met DNA. Daarom worden ze "indirect" genoemd.
De eerste die beschreef was de trans-activator van het virus van "Herpes eenvoudig " (HSV) bekend als VP16, die bindt aan de OCT-1-factor wanneer de cellen zijn geïnfecteerd met dit virus, waardoor de transcriptie van een specifiek gen wordt gestimuleerd.
De factoren van dit type, zoals die die bindend zijn aan het DNA, kunnen gentranscriptie activeren of onderdrukken, zodat ze respectievelijk "dwangers" en "gecorrigeerd" worden genoemd.
Regulatie
Deze eiwitten kunnen op twee niveaus worden gereguleerd: in hun synthese en in hun activiteit, die afhankelijk is van verschillende variabelen en meerdere situaties.
Synthesegeling
De regulatie van de synthese kan gerelateerd zijn aan het expressieweefselspecifieke van bepaalde transcriptiefactoren. Een voorbeeld hiervan kan de myod -factor zijn, alleen gesynthetiseerd in skeletspiercellen en is noodzakelijk voor de differentiatie van zijn ongedifferentieerde fibroblasten.
Kan u van dienst zijn: gekoppelde genenHoewel de regulatie van synthese fundamenteel wordt gebruikt voor de controle van genetische expressie in specifieke celtypen en weefsels, is dit niet de enige manier, omdat de synthese van factoren die betrokken zijn bij de inductie van genen die deelnemen aan de respons ook wordt gereguleerd naar verschillende stimuli.
Activiteitsverordening
Een ander regulatiemechanisme voor transcriptiefactoren is de regulatie van de activiteit ervan, wat te maken heeft met de activering van andere pre -bestaande transcriptiefactoren die positieve of negatieve effecten hebben op de activiteit van een bepaalde factor.
De activering van deze "secundaire" factoren treedt meestal op door verschillende mechanismen zoals ligandunie, veranderingen in eiwit-eiwit, fosforyleringsinteracties, onder andere.
Functies en belang
Transcriptiefactoren nemen deel aan verschillende processen zoals embryonale ontwikkeling, groei en differentiatie, celcycluscontrole, aanpassing aan fluctuerende omgevingscondities, het behoud van specifieke eiwitsynthesepatronen van cellen en weefsels, enz.
In planten hebben ze bijvoorbeeld belangrijke functies in de verdediging en in reactie -gebeurtenissen op verschillende soorten stress. Er is vastgesteld dat osteogenese bij dieren wordt geregeld door transcriptiefactoren, evenals vele andere differentiatieprocessen van verschillende cellijnen.
Gezien het belang van deze eiwitten in organismen, is het niet ongewoon om te denken dat veranderingen in deze regelgevende elementen ernstige pathologische veranderingen zullen veroorzaken.
In het geval van mensen kunnen pathologieën geassocieerd met transcriptiefactoren ontwikkelingsstoornissen zijn (vanwege mutaties die bijvoorbeeld inactivering van transcriptiefactoren produceren), aandoeningen in de hormonale respons of kankers.
Referenties
- Alberts, B., Dennis, B., Hopkin, K., Johnson, a., Lewis, J., Raff, m.,... Walter, P. (2004). Essentiële celbiologie. Abingdon: Garland Science, Taylor & Francis Group.
- Iwafuchi-doi, m., & Zaret, K. S. (2014). Pioneer transcriptiefactoren bij het herprogrammeren van cellen. Genen en ontwikkeling, 28, 2679-2692.
- Latchman, D. (1997). Transcriptiefactoren: een overzicht. Inteken. J. Biochem. Cel. Biol., 29(12), 1305-1312.
- Latchman, D. S. (2007). Transcriptiefactoren. Encyclopedia of Life Sciences, 1-5.
- Marie, p. J. (2008). Transcriptiefactoren die osteoblastogenese regelen. Archieven van biochemie en biofysica, 473, 98-105.
- Pabo, c., & Sauer, r. T. (1992). Transcriptiefactoren: structurele families en principes van DNA -herkenning. Annu. Revisie., 61, 1053-1095.
- Singh, K. B., Foley, r. C., & Oñate-sánchez, l. (2002). Transcriptiefactoren in plantenverdediging en stressrespons. Huidige mening in de plantenbiologie, 5, 430-436.
- « Buskenmerken, voedsel, reproductie, habitat
- Chlorofytes kenmerken, habitat, reproductie, voedsel »